
Piras, I. S., Vacca, L., Calò, C. M., Falchi,
A., Ghiani, M. E., Varesi, L., Vona, G., 2005, Distribuzione delle varianti  32 e 64I dei
geni per il CCR5 e il CCR2 in alcune popolazioni del Mediterraneo. Antropo, 9,
29-39. www.didac.ehu.es/antropo
32 e 64I dei
geni per il CCR5 e il CCR2 in alcune popolazioni del Mediterraneo. Antropo, 9,
29-39. www.didac.ehu.es/antropo
Distribuzione
delle varianti 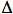 32 e 64I dei geni per il CCR5 e il CCR2 in alcune
popolazioni del Mediterraneo
32 e 64I dei geni per il CCR5 e il CCR2 in alcune
popolazioni del Mediterraneo
Distribution of 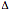 32 and 64I variants
of CCR5 and CCR2 genes in some Mediterranean populations
32 and 64I variants
of CCR5 and CCR2 genes in some Mediterranean populations
Ignazio
Stefano Piras1, Lucia Vacca1, Carla Maria Calò1,
Alessandra Falchi1,2, Maria Elena Ghiani1, Laurent Varesi2,
Giuseppe Vona1
1
Dip. di Biologia Sperimentale, Università di Cagliari (Italia)
2 Faculty of Sciences and Techniques,
University of Corse (France)
Per
la corrispondenza: Ignazio Stefano Piras. Università di Cagliari. Dipartimento
di Biologia Sperimentale, Sez. Scienze Antropologiche. SS 554, km 4,500 – 09042
Monserrato (Ca) – Italia. E-mail: ispiras@unica.it
Parole
chiave:
CCR5, CCR2, HIV-1, popolazioni Mediterranee
Key words:
CCR5, CCR2, HIV-1, Mediterranean populations
Riassunto
Le
varianti dei geni CCR5 e CCR2 sono state messe in relazione con una differente
suscettibilità all’infezione da HIV-1. In particolare, gli individui omozigoti
per il CCR5-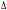 32 mostrano una protezione quasi completa all’infezione,
mentre gli individui eterozigoti presentano una progressione ritardata della
sindrome. Gli individui che possiedono la mutazione CCR2-64I presentano invece
una ritardata progressione della sindrome sia nella condizione omozigote che
nella condizione eterozigote. In
questo studio sono stati analizzati i due polimorfismi in alcune popolazioni
del Mediterraneo.
32 mostrano una protezione quasi completa all’infezione,
mentre gli individui eterozigoti presentano una progressione ritardata della
sindrome. Gli individui che possiedono la mutazione CCR2-64I presentano invece
una ritardata progressione della sindrome sia nella condizione omozigote che
nella condizione eterozigote. In
questo studio sono stati analizzati i due polimorfismi in alcune popolazioni
del Mediterraneo.
I
risultati confermano la presenza di queste varianti nelle popolazioni del
Mediterraneo, e sembrano indicare la validità di questi marcatori per l’analisi
popolazionistica.
Abstract
Variants of CCR5 and CCR2 genes are related with different
susceptibility to HIV-1 infection. Particulary, homozygous subjects for CCR5-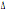 32 mutation show an high
protection against infection, while heterozygous subjects show a delayed
disease progression. Both homozygous and heterozygous subjects carrying
CCR2-64I mutation show delayed disease progression.
32 mutation show an high
protection against infection, while heterozygous subjects show a delayed
disease progression. Both homozygous and heterozygous subjects carrying
CCR2-64I mutation show delayed disease progression.
In this study we analyse these polymorphisms
in some Mediterranean populations. Results confirm the presence of these
variants in Mediterranean populations and suggest the validity of these markers
for populations genetic analysis.
Introduzione
Alcune varianti dei geni codificanti per i
recettori CCR5 e CCR2 sono stati messi in relazione con una diversa
suscettibilità all’infezione da HIV–1. I due geni sono situati sul cromosoma 3 in posizione 3p21.3-p24,
all’interno di un cluster che include la maggior parte dei geni codificanti per
i recettori delle chemochine (Samson et al., 1996). Il CCR5 è espresso sulla
superficie della membrana dei monociti, dei macrofagi, delle cellule T della memoria e sulle cellule
dendritiche; oltre a legare le beta chemochine RANTES, MIP-1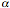 e MIP-1
e MIP-1 il CCR5 è un
importante corecettore per l’HIV–1 (Samson et al., 1996; Huang Y et al., 1996).
il CCR5 è un
importante corecettore per l’HIV–1 (Samson et al., 1996; Huang Y et al., 1996).
I soggetti omozigoti per
una delezione di 32 bp (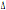 32) nel segmento del gene CCR5 che codifica per il secondo dominio
extracellulare della proteina sono
resistenti all’infezione (Aarons et al., 1997; Samson et al., 1996; Dean et
al., 1996; Liu et al., 1996; Biti et al., 1997; Rana et al., 1997; Huang et
al., 1996) anche se tale protezione non è totalmente completa (Biti et al.,1997; O’Brien et al., 1997; Theodorou
et al., 1997). Gli eterozigoti appaiono parzialmente protetti dall’infezione
(Samson M., et al., 1996; Michael et al., 1997) e mostrano un rallentamento nella
progressione della sindrome (Huang et al., 1996; Rappaport et al., 1997;
Eugen-Olsen et al., 1997; Michael
et al.,1997). La distribuzione del CCR5-
32) nel segmento del gene CCR5 che codifica per il secondo dominio
extracellulare della proteina sono
resistenti all’infezione (Aarons et al., 1997; Samson et al., 1996; Dean et
al., 1996; Liu et al., 1996; Biti et al., 1997; Rana et al., 1997; Huang et
al., 1996) anche se tale protezione non è totalmente completa (Biti et al.,1997; O’Brien et al., 1997; Theodorou
et al., 1997). Gli eterozigoti appaiono parzialmente protetti dall’infezione
(Samson M., et al., 1996; Michael et al., 1997) e mostrano un rallentamento nella
progressione della sindrome (Huang et al., 1996; Rappaport et al., 1997;
Eugen-Olsen et al., 1997; Michael
et al.,1997). La distribuzione del CCR5-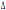 32 è limitata prevalentemente all’Europa e al Nord Africa a parte alcune
eccezioni (Martinson et al., 1997; Su et al., 1998; Su et al., 2000; Smith et
al., 1997; Barbouche et al., 2001), mostrando un gradiente Nord - Sud, con le
maggiori frequenze nelle popolazioni Finlandese e Mordoviniana (16%) e minori
in Sardegna (4%) (Libert et al., 1998). Tale distribuzione è stata spiegata con
un’origine recente della mutazione nel Nord – Est Europa, datata circa 700 anni
fa (con un range compreso tra 275 – 1870 anni) (Stephens et al., 1998; Libert
et al., 1998). Il vantaggio selettivo che conferiva nei confronti del vaiolo
avrebbe permesso alla mutazione di mantenere elevate frequenze (Galvani et al.,
2003), mentre la presenza nel sud - Europa e il gradiente Nord - Sud sono stati spiegati con l’ipotesi di una
dispersione dell’allele da parte dei Vichinghi durante i loro spostamenti dal
loro luogo d’origine nel nord Europa alle altre regioni (Lucotte, 2003).
32 è limitata prevalentemente all’Europa e al Nord Africa a parte alcune
eccezioni (Martinson et al., 1997; Su et al., 1998; Su et al., 2000; Smith et
al., 1997; Barbouche et al., 2001), mostrando un gradiente Nord - Sud, con le
maggiori frequenze nelle popolazioni Finlandese e Mordoviniana (16%) e minori
in Sardegna (4%) (Libert et al., 1998). Tale distribuzione è stata spiegata con
un’origine recente della mutazione nel Nord – Est Europa, datata circa 700 anni
fa (con un range compreso tra 275 – 1870 anni) (Stephens et al., 1998; Libert
et al., 1998). Il vantaggio selettivo che conferiva nei confronti del vaiolo
avrebbe permesso alla mutazione di mantenere elevate frequenze (Galvani et al.,
2003), mentre la presenza nel sud - Europa e il gradiente Nord - Sud sono stati spiegati con l’ipotesi di una
dispersione dell’allele da parte dei Vichinghi durante i loro spostamenti dal
loro luogo d’origine nel nord Europa alle altre regioni (Lucotte, 2003).
Il CCR2 è il recettore
per le chemochine MCP–1,2,3,4 e 5 (D’Souza et al., 1996) e anch’esso è un
corecettore per l’HIV–1. Una sostituzione 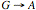 in
posizione 190 determina la sostituzione di una valina con una isoleucina nel
primo dominio transmembrana del recettore. Nei portatori di questa mutazione è
stata osservato una rallentamento nella progressione della sindrome ma non una protezione dalla
trasmissione da HIV-1 (Smith et
al., 1997; Michael et al., 1997). Il CCR2-64I possiede un’origine precedente
al CCR5-
in
posizione 190 determina la sostituzione di una valina con una isoleucina nel
primo dominio transmembrana del recettore. Nei portatori di questa mutazione è
stata osservato una rallentamento nella progressione della sindrome ma non una protezione dalla
trasmissione da HIV-1 (Smith et
al., 1997; Michael et al., 1997). Il CCR2-64I possiede un’origine precedente
al CCR5-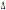 32 (circa 32000 anni) (Kostrikis and Martinson, 1999), ed è presente in
tutti i continenti, raggiungendo le frequenze più elevate (sino al 57%) nelle
popolazioni asiatiche (Martinson et al., 2000; Struyf et al., 2000; Su et al.,
1999; Su et al., 2000).
32 (circa 32000 anni) (Kostrikis and Martinson, 1999), ed è presente in
tutti i continenti, raggiungendo le frequenze più elevate (sino al 57%) nelle
popolazioni asiatiche (Martinson et al., 2000; Struyf et al., 2000; Su et al.,
1999; Su et al., 2000).
In questo lavoro si è
studiata la distribuzione di questi due polimorfismi in quattro popolazioni del
bacino del Mediterraneo (Sardegna, Corsica, Isole Baleari e Turchia), data
l’importanza che tali marcatori possiedono nella trasmissione dell’HIV–1 e
considerato che, soprattutto per il CCR2–64I, non sono presenti molti dati in
letteratura relativi a queste popolazioni. Inoltre, attraverso i confronti con
i dati rinvenibili in letteratura, si è analizzata la validità di tali
marcatori per gli studi popolazionistici.
Materiali e Metodi
È stato esaminato un
campione di 315 individui sani non imparentati tra loro provenienti da diverse
aree del Mediterraneo (Sardegna, Corsica, Turchia e Baleari). Il DNA genomico è
stato estratto da sangue intero con la metodica del fenolo – cloroformio e successivamente è stato amplificato
mediante PCR secondo le condizioni descritte da altri autori (de Roda Husman et
al., 1997; Struyf et al., 1999). Per il CCR5 la determinazione dei genotipi è
stata effettuata su gel di agarosio al 4%. Per l’allele mutato il frammento
ottenuto era di 161 bp mentre per l’allele selvatico era di 193 bp. Per il CCR2
dopo la PCR è stato necessario eseguire una digestione con l’enzima di
restrizione BstF5 I (4 ore a 65° C) e una elettroforesi su gel di agarosio al
4%. I frammenti ottenuti dopo la digestione erano: 360 bp, 203 bp e 102 bp per
il genotipo wt/wt; 360, 203, 160 e 102 bp per il genotipo wt/64I; 203, 160 e
102 bp per il genotipo 64I/64I.
Dai dati ottenuti sono
state calcolate le frequenze alleliche, genotipiche, il test di Fisher basato
sulle frequenze genotipiche con il programma Genepop 1.2 (Raymond and Rousset
1995); per valutare se il campione fosse in equilibrio di Hardy Weinberg è
stato applicato il test statistico della catena di Markov (Guo e Thompson
1992), mentre le frequenze aplotipiche sono state stimate con la procedura
Maximum-Likelihood usando l’algoritmo expectation-maximation (EM) (Excoffier
and Slatkin, 1995). È stata testata l’esistenza di un Linkage Disequilibrium
tra i due loci ed è stato effettuato il Test di Differenziazione di Fisher
basato sulle frequenze aplotipiche. Questi parametri sono stati calcolati con
il software Arlequin 2000 (Schneider et al., 2000). Per valutare l’eventuale
causa del Linkage Disequilibrium è stato effettuato il Test di Otha (Ohta et
al., 1969) con il programma Genetix v 4.03 (Belkhir et al., 2002).
Utilizzando i dati
disponibili in letteratura e con l’ausilio del software R-Matrix (Harpending
and Jenkins 1973) sono stati effettuati dei confronti con le altre popolazioni.
Nel tentativo di spiegare
la differenziazione delle popolazioni tenendo conto della loro localizzazione
geografica è stato utilizzato l’indice di autocorrelazione spaziale I di
Moran. Il calcolo del coefficiente
viene effettuato raggruppando le popolazioni in classi di distanze
geograficamente arbitrarie. L’Indice di Moran varia tra +1 e -1. Il suo valore
atteso in assenza di autocorrelazione si approssima a zero. Un valore positivo
indica similarità genetica nella classe di distanze analizzate, un valore
negativo indica una dissimilarità nelle frequenze dell’allele. Il
correlogramma, che riporta i coefficienti di autocorrelazione rispetto alle
distanze, descrive l’andamento nello spazio della variazione genetica mostrata
per ciascun allele. I correlogrammi sono riconducibili ad alcuni andamenti
tipici, ognuno dei quali descrive l’affinità o la diversità genetica delle
popolazioni in funzione della localizzazione geografica. L’analisi è stata
effettuata con il programma SAAP 4.3
Risultati
Le frequenze alleliche, genotipiche e
aplotipiche sono mostrate in tabella 1. I risultati indicano per il CCR5-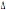 32 dei
valori compresi tra 8,9% delle Isole Baleari e l’1,6% della Sardegna, mentre
per il CCR2 64I abbiamo dei valori compresi tra l’11% della Turchia e il 6,8%
della Sardegna.
32 dei
valori compresi tra 8,9% delle Isole Baleari e l’1,6% della Sardegna, mentre
per il CCR2 64I abbiamo dei valori compresi tra l’11% della Turchia e il 6,8%
della Sardegna.
Per quanto riguarda le
frequenze genotipiche, per il locus CCR5 si può evidenziare la presenza del
genotipo 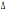 32/
32/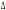 32 solamente nel campione delle Isole Baleari. Per il locus CCR2 si può
notare l’assenza in tutte le popolazioni del genotipo 64I/64I. Per tutti e due i loci il genotipo più
frequente è il genotipo selvatico wt/wt.
32 solamente nel campione delle Isole Baleari. Per il locus CCR2 si può
notare l’assenza in tutte le popolazioni del genotipo 64I/64I. Per tutti e due i loci il genotipo più
frequente è il genotipo selvatico wt/wt.
Sono stati individuati tutti gli aplotipi
possibili e in tutte le popolazioni l’aplotipo più frequente è quello
costituito dai due alleli selvatici (wt/wt), mentre il meno frequente è quello
costituito dai due alleli mutati 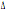 32/64I. Questo aplotipo è presente
nella Turchia e nelle Isole Baleari rispettivamente con 1,9% e 0,2%, risultando
assente nel campione della Sardegna e della Corsica.
32/64I. Questo aplotipo è presente
nella Turchia e nelle Isole Baleari rispettivamente con 1,9% e 0,2%, risultando
assente nel campione della Sardegna e della Corsica.
|
|
Frequenze alleliche
|
Frequenze genotipiche
|
Frequenze aplotipiche
|
|
|
CCR5
|
CCR2
|
CCR5
|
CCR2
|
|
|
wt
|
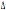 32 32
|
wt
|
64I
|
wt/
wt
|
wt/
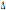 32 32
|
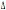 32/ 32/
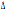 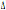 32 32
|
wt/
wt
|
wt/
64I
|
64I/
64I
|
wt/
wt
|
wt/
64I
|
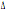 32/ 32/
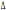 wt wt
|
 32/ 32/
 64I 64I
|
|
Cor
|
94,8
|
5,2
|
92,8
|
7,2
|
89,6
|
10,4
|
0,0
|
87,2
|
12,8
|
0,0
|
87,6
|
7,2
|
5,2
|
0,0
|
|
Sar
|
98,4
|
1,6
|
93,2
|
6,8
|
96,8
|
3,2
|
0,0
|
86,3
|
13,7
|
0,0
|
91,6
|
6,8
|
1,6
|
0,0
|
|
Tur
|
97,0
|
3,0
|
89,0
|
11,0
|
94,0
|
6,0
|
0,0
|
78,0
|
22,0
|
0,0
|
87,9
|
9,1
|
1,1
|
1,9
|
|
Bal
|
91,1
|
8,9
|
91,1
|
8,9
|
84,5
|
13,3
|
0,0
|
82,2
|
17,8
|
0,0
|
82,4
|
8,7
|
8,7
|
0,2
|
Tabella 1. Frequenze alleliche, genotipiche e aplotipiche per
le popolazioni studiate.
Table 1. Allele, genotype and haplotype frequencies for the
studied populations.
Per valutare la differenziazione tra le
popolazioni è stato applicato il Test esatto di Fisher, sia sulle frequenze
genotipiche sia su quelle aplotipiche.
Le popolazioni in base al test esatto di Fisher
appaiono differenziate tra di loro a livello genotipico solamente per il locus
CCR5 (P = 0,02772 ± 0,00329) e non per
il CCR2 (P = 0,05604 ± 0,01201).
Effettuando il confronto a coppie tra popolazioni per ogni locus l’unico valore
significativo riscontrato è stato quello tra la Sardegna e le Baleari per il
locus CCR5 (P = 0,0054 ± 0,00062)
A livello aplotipico le popolazioni non risultano
differenziate né a livello complessivo (P = 0,0637 ± 0,04357) né nei confronti a coppie.
È stato riscontrato un valore significativo di
Linkage Disequilibrium per il campione proveniente dalla Turchia (P = 0,01173 ± 0,00304). Il test di Otha suggerisce la deriva genica come causa del
Linkage.
Per verificare l’esistenza di una correlazione
spaziale per gli alleli analizzati tra le popolazioni prese in esame e quelle
studiate da altri autori per gli stessi loci, è stato calcolato l’Indice di
Moran (I). Tale indice è risultato significativo per il locus CCR2 il cui
correlogramma (figura 1) costruito sui coefficienti di correlazione mostra un
andamento assimilabile a un gradiente.
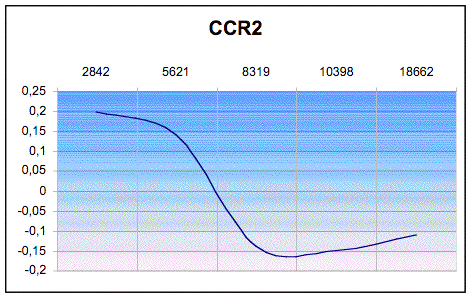
Figura 1. Correlogramma per il locus CCR2.
Figure 1. Correlogram for CCR2 locus.
I nostri dati sono stati confrontati con quelli
presenti in letteratura (tabella 2; figura 2). Per quanto riguarda l’Europa e
il Mediterraneo dalla figura 2 si può notare che il range di variazione degli
alleli dei loci esaminati sono: per il CCR5- 32 le frequenze sono comprese tra il 14% (Polonia) e l’1,6%
(Sardegna); per il CCR2-64I le frequenze variano tra il 18% (Polonia) e il 6,8%
(Sardegna). Si è poi operato un confronto con le popolazioni di altri
continenti utilizzando le frequenze alleliche mediante l’analisi dell’R-matrix
(Harpending and Jenkins 1973). Dal grafico costruito utilizzando le prime due
componenti (figura 3) si può notare che la prima componente, che rappresenta il
68% della variabilità totale, separa le popolazioni europee (che hanno valori
positivi) dalle altre popolazioni. La seconda componente (32% di variabilità)
separa invece le popolazioni nord europee da quelle del sud Europa, e quelle
asiatiche da quelle africane, che clusterizzano entrambe in un quadrante a
parte. La popolazione che ha contribuito maggiormente alla variabilità
riscontrata è la Polonia, mentre quella che ha contribuito in minor misura è
l’Italia.
32 le frequenze sono comprese tra il 14% (Polonia) e l’1,6%
(Sardegna); per il CCR2-64I le frequenze variano tra il 18% (Polonia) e il 6,8%
(Sardegna). Si è poi operato un confronto con le popolazioni di altri
continenti utilizzando le frequenze alleliche mediante l’analisi dell’R-matrix
(Harpending and Jenkins 1973). Dal grafico costruito utilizzando le prime due
componenti (figura 3) si può notare che la prima componente, che rappresenta il
68% della variabilità totale, separa le popolazioni europee (che hanno valori
positivi) dalle altre popolazioni. La seconda componente (32% di variabilità)
separa invece le popolazioni nord europee da quelle del sud Europa, e quelle
asiatiche da quelle africane, che clusterizzano entrambe in un quadrante a
parte. La popolazione che ha contribuito maggiormente alla variabilità
riscontrata è la Polonia, mentre quella che ha contribuito in minor misura è
l’Italia.
|
Europa
|
D32
|
64I
|
Rif. Bibliografici
|
|
Corsica
|
5,2
|
7,2
|
Presente studio
|
|
Sardegna
|
1,6
|
6,8
|
Presente studio
|
|
Turchia
|
3,0
|
11,0
|
Presente studio
|
|
Baleari
|
8,9
|
8,9
|
Presente studio
|
|
Italia
|
3,0
|
13,0
|
Su et al., 2000
|
|
Nord Europa
|
9,0
|
16,0
|
Su et al., 2000
|
|
Germania
|
7,0
|
13,0
|
Su et al., 2000
|
|
Spagna
|
2,0
|
8,0
|
Su et al., 2000
|
|
Polonia
|
14,0
|
18,0
|
Su et al., 2000
|
|
Belgio
|
11,9
|
7,4
|
Struyf et al., 2000
|
|
Grecia
|
5,2
|
14,6
|
Papa et al., 2000
|
|
Africa
|
D32
|
64I
|
|
|
Pigmei Biaka
|
0,0
|
11,0
|
Su et al., 2000
|
|
Pigmei Mbuti
|
0,0
|
10,0
|
Su et al., 2000
|
|
Lissongo
|
0,0
|
14,0
|
Su et al., 2000
|
|
Benin
|
0,0
|
11,0
|
Su et al., 2000
|
|
Sudan
|
0,0
|
19,0
|
Su et al., 2000
|
|
Nigeria
|
0,0
|
12,0
|
Su et al., 2000
|
|
Tunisia
|
1,0
|
19,3
|
Barbouche et al., 2001
|
|
America
|
D32
|
64I
|
|
|
Karitiana
|
0,0
|
7,0
|
Su et al., 2000
|
|
Surui
|
0,0
|
3,0
|
Su et al., 2000
|
|
Mayan
|
0,0
|
33,0
|
Su et al., 2000
|
|
Nativi Americani
|
0,0
|
31,1
|
Iyer et al., 2001
|
|
Asia - Oceania
|
D32
|
64I
|
|
|
Aborigeni Australiani
|
0,0
|
10,0
|
Su et al., 2000
|
|
Nuova Guinea
|
0,0
|
14,0
|
Su et al., 2000
|
|
Nord Cina
|
0,0
|
20,0
|
Su et al., 2000
|
|
Est Cina
|
0,0
|
19,2
|
Su et al., 2000
|
|
Tibet
|
2,0
|
28,0
|
Su et al., 2000
|
|
Nord Est Thailandia
|
0,0
|
17,0
|
Su et al., 2000
|
|
Malesia
|
0,0
|
22,0
|
Su et al., 2000
|
|
Giava
|
0,0
|
10,0
|
Su et al., 2000
|
|
Cambogia
|
1,0
|
18,5
|
Su et al., 2000
|
|
Corea
|
0,0
|
26,0
|
Su et al., 2000
|
|
Giappone
|
0,0
|
17,5
|
Su et al., 2000
|
|
Buryat
|
0,0
|
57,0
|
Su et al., 2000
|
|
Assam caste
|
0,0
|
22,0
|
Su et al., 2000
|
|
Rajasthani
|
5,0
|
5,0
|
Su et al., 2000
|
|
Ahom
|
0,0
|
19,0
|
Su et al., 2000
|
|
Sud Iran
|
1,5
|
12,2
|
Gharagozloo et al., 2005
|
|
Emirati Arabi
|
0,0
|
6,0
|
Su et al., 2000
|
Tabella 2. Frequenze alleliche dei
marcatori analizzati in popolazioni provenienti dai diversi continenti.
Table 2. Allele frequencies for
the used genetic markers in populations from different continents.
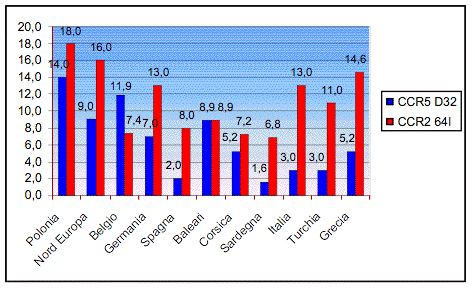
Figura 2. Distribuzione del CCR5- 32 e del
CCR2 64I in Europa.
32 e del
CCR2 64I in Europa.
Figure 2. CCR5- 32 and
CCR2-64I distribution in Europe.
32 and
CCR2-64I distribution in Europe.
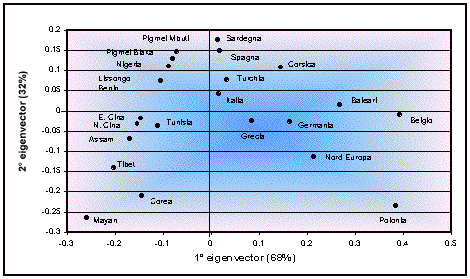
Figura 3. Analisi
dell’r-matrix: plot dei primi due eigenvectors.
Figure 3. R-matrix analysis: plot of the first two
eigenvectors.
Discussione
In questo lavoro è stata
studiata la distribuzione dei polimorfismi CCR5-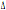 32 e CCR2-64I in alcune popolazioni provenienti dalle principali isole del
Mediterraneo (Isole Baleari, Sardegna e Corsica) e dalla Turchia. I risultati
mostrano che le frequenze alleliche del CCR5-
32 e CCR2-64I in alcune popolazioni provenienti dalle principali isole del
Mediterraneo (Isole Baleari, Sardegna e Corsica) e dalla Turchia. I risultati
mostrano che le frequenze alleliche del CCR5-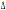 32 hanno dei valori che vanno dal 8,9% delle Isole Baleari all’1,6% della
Sardegna. Il valore riscontrato per la Sardegna si è inferiore a quello
riportato da Libert et al. (1998), che riportano un 4%, mentre si discosta poco
dalle frequenze indicate da Battiloro et al. (2000) (2,1%). Per la Corsica
(Ajaccio) Lucotte and Mercier (1998) riportano un 0,9%, contro un 5,2% del
nostro campione. Tale differenza può essere dovuta al motivo che il campione da
noi esaminato proveniva da diverse aree isolate della Corsica come Corte e
Balagna. La Turchia nella nostra analisi mostra un valore del 3%, simile a
quello riportato da Degerli et al. 2005 (2,18%) e inferiore a quello riportato
da Libert et al., 1998 (6,3%). Per quanto riguarda le Isole Baleari è stato
osservato un valore del
32 hanno dei valori che vanno dal 8,9% delle Isole Baleari all’1,6% della
Sardegna. Il valore riscontrato per la Sardegna si è inferiore a quello
riportato da Libert et al. (1998), che riportano un 4%, mentre si discosta poco
dalle frequenze indicate da Battiloro et al. (2000) (2,1%). Per la Corsica
(Ajaccio) Lucotte and Mercier (1998) riportano un 0,9%, contro un 5,2% del
nostro campione. Tale differenza può essere dovuta al motivo che il campione da
noi esaminato proveniva da diverse aree isolate della Corsica come Corte e
Balagna. La Turchia nella nostra analisi mostra un valore del 3%, simile a
quello riportato da Degerli et al. 2005 (2,18%) e inferiore a quello riportato
da Libert et al., 1998 (6,3%). Per quanto riguarda le Isole Baleari è stato
osservato un valore del 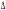 32 del 8,9%. In generale le frequenze riportate nel nostro lavoro hanno dei
valori nell’ordine di quelli osservati da altri autori per le popolazioni
dell’Europa, dove è presente un gradiente Nord – Sud, con i valori inferiori
presenti nelle regioni meridionali (Libert et al., 1998). Solo le Isole Baleari
sembrano possedere una frequenza più elevata, come riportato
da Alvarez et al. (1998) per altre popolazioni iberiche, le quali mostrano
frequenze del
32 del 8,9%. In generale le frequenze riportate nel nostro lavoro hanno dei
valori nell’ordine di quelli osservati da altri autori per le popolazioni
dell’Europa, dove è presente un gradiente Nord – Sud, con i valori inferiori
presenti nelle regioni meridionali (Libert et al., 1998). Solo le Isole Baleari
sembrano possedere una frequenza più elevata, come riportato
da Alvarez et al. (1998) per altre popolazioni iberiche, le quali mostrano
frequenze del 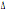 32 che arrivano sino al 9,6%.
32 che arrivano sino al 9,6%.
Per quanto riguarda il
CCR2-64I, la Sardegna e la Corsica sono caratterizzate da valori che si posizionano
fra i più bassi rispetto a quelli sinora riportati per le popolazioni europee.
La frequenza è paragonabile solo a quella del Belgio, con un 7,4% (Struyf et
al., 1999).
In generale la
distribuzione del CCR5-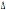 32 è limitata all’Europa e al Nord Africa a parte alcune eccezioni, che
riguardano il Tibet con un 2% e il Rajasthan con il 5% (Martinson et al., 1997;
Su et al., 1998; Su et al., 2000; Smith et al., 1997; Barbouche et al., 2001)
(tabella 2).
32 è limitata all’Europa e al Nord Africa a parte alcune eccezioni, che
riguardano il Tibet con un 2% e il Rajasthan con il 5% (Martinson et al., 1997;
Su et al., 1998; Su et al., 2000; Smith et al., 1997; Barbouche et al., 2001)
(tabella 2).
Il CCR2-64I possiede
un’origine più antica del CCR5-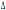 32 risalendo a circa 32000 anni (Kostrikis and Martinson, 1999), ciò è
testimoniato dalla sua presenza in tutti i continenti. In particolare le
frequenze più elevate sono state riscontrate tra i Buriati (popolazione che
vive nella Siberia orientale, al confine con la Mongolia), dove è stata
osservata una frequenza del 57% e tra alcune popolazioni native americane come
i Mayan (33%). Tra le altre popolazioni asiatiche la frequenza del 64I è compresa tra il 28% e il 6%.
Nel continente africano i valori sono compresi tra il 19,3% e il 10% mentre in
Europa sono compresi tra il 18% e il 6,8% (Martinson et al., 2000; Struyf et
al., 2000; Su et al., 1999; Su et al., 2000; Barbouche et al., 2001).
32 risalendo a circa 32000 anni (Kostrikis and Martinson, 1999), ciò è
testimoniato dalla sua presenza in tutti i continenti. In particolare le
frequenze più elevate sono state riscontrate tra i Buriati (popolazione che
vive nella Siberia orientale, al confine con la Mongolia), dove è stata
osservata una frequenza del 57% e tra alcune popolazioni native americane come
i Mayan (33%). Tra le altre popolazioni asiatiche la frequenza del 64I è compresa tra il 28% e il 6%.
Nel continente africano i valori sono compresi tra il 19,3% e il 10% mentre in
Europa sono compresi tra il 18% e il 6,8% (Martinson et al., 2000; Struyf et
al., 2000; Su et al., 1999; Su et al., 2000; Barbouche et al., 2001).
L’analisi dell’R-matrix rivela
una separazione abbastanza puntuale tra le popolazioni dei tre continenti
Africa, Asia ed Europa. Il grafico
ricavato dall’analisi dell R-matrix (figura 3) raggruppa nel quadrante con i
valori positivi dei due eigenvector alcune popolazioni dell’area mediterranea
che hanno avuto relazioni più o meno strette tra loro. Manca in questo cluster
la Grecia che ha avuto un forte peso sulla composizione genetica dell’Italia e
della Corsica (Piazza et al., 1988).
Sardegna e Spagna
appaiono le popolazioni più affini. È nota la presenza spagnola nell’isola
protrattasi per diversi secoli che sicuramente può aver determinato tale
affinità già segnalata da precedenti lavori (Vona et al., 1994). Ma influssi
dalla penisola iberica sulla Sardegna erano presenti già in epoca preistorica
come indicato da lavori sul mtDNA e sugli aplotipi del cromosoma Y (Torroni et
al., 1998; Scozzari et al., 2001; Tofanelli et al., 2004).
I dati provenienti dai
marcatori classici e dal DNA mitocondriale suggeriscono un’origine comune delle
popolazioni delle due isole e quindi una certa similarità genetica (Varesi et
al., 2000; Vona et al., 2002; Vona et al., 2003). I risultati confermano
l’affinità tra corsi e sardi. La non perfetta identità tra le due isole,
sarebbe da attribuire alla loro storia demografica (bassa densità di
popolazione, sottopopolazioni isolate, epidemie), che avrebbe portato a fenomeni di deriva genetica e
all’isolamento (Vona, 1997; Vona et al., 2002).
La presenza dell’Italia
continentale nello stesso cluster con la Sardegna e la Corsica sembra indicare
che le due isole hanno avuto apporti continentali. Questo fatto è dimostrato da
altre analisi sui marcatori aploidi del mtDNA e dell’Y (Morelli et al., 2000;
Tofanelli et al., 2004).
La particolare posizione
delle Isole Baleari all’interno del cluster può trovare una possibile
spiegazione nella peculiarità genetica della popolazione di Ibiza. Per alcuni
marcatori tale isola sarebbe più simile alle popolazioni del Nord Africa
(Picornell et al., 1996) che alle altre due isole dell’arcipelago (Maiorca e
Minorca) e alle altre popolazioni del Mediterraneo (Moral 1986; Miguel and
PetitPierre 1989; Miguel et al., 1990; Massanet et al., 1997).
Le frequenze relative
all’allele CCR2-64I sembrano indicare un gradiente dell’allele dal centro
europa, dove presenta la sua massima frequenza verso due direzioni: il
Mediterraneo occidentale e quello orientale. Lo stesso andamento si può notare
per l’allele CCR5-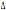 32. Questi risultati sembrano convalidare le osservazioni riportate da Libert et al. (1998) per il CCR5-
32. Questi risultati sembrano convalidare le osservazioni riportate da Libert et al. (1998) per il CCR5-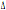 32 sull’esistenza di un gradiente e l’andamento geografico espresso dal
correlogramma per il locus CCR2 (figura 1).
32 sull’esistenza di un gradiente e l’andamento geografico espresso dal
correlogramma per il locus CCR2 (figura 1).
In generale dai risultati
ottenuti si può affermare che i due marcatori hanno mostrato una certa validità
dell’impiego dei marcatori nelle analisi popolazionistiche sia a livello delle
popolazioni continentali sia a livello delle popolazioni intracontinentali.
Bibliografia
Aarons, E., Fernandez, M., Rees, A., McClure, M., e Weber, J., 1997,
CC-chemokine receptor 5 genotypes and in vitro susceptibility to HIV-1 of a
cohort of British HIV-exposed uninfected homosexual men. AIDS, 11(5):688-689.
Alvarez, V.,
Lopez-Larrea, C., e Coto, E., 1998, Mutational analysis of the CCR5 and CXCR4
genes (HIV-1 co-receptors) in resistance to HIV-1 infection and AIDS
development among intravenous drug users. Hum Genet, 102(4), 483-486.
Barbouche, R.M.,
Hong, L., Dellagi, K., e Kostrikis, L.G., 2001, Contrasting frequencies of
CCR5delta32 and CCR2-64I alleles in the Tunisian population. J Acquir Immune Defic
Syndr, 26(3), 298-299.
Battiloro, E., Androni, M., Parisi, S.G., Mura,
M.S., Sotgiu, G., Aceti, A., Cossu, G., Concia, E., Verna, R., e D'Ambrosio,
E., 2000, Distribution of the CCR5 delta32 allele in Italian HIV type
1-infected and normal individuals. AIDS Res Hum Retroviruses, 16(2), 181-182.
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N., e
Bonhomme, F., 2002, genetix 4.03, logiciel sous
Windows TM pour la génétique des populations. Laboratoire Génome, Populations,
Interactions, CNRS UMR 5000, Université de Montpellier II, Montpellier, France.
Biti, R.,
French, R., Young, J., Bennet, B., e Steward, G., 1997, HIV-1 infection in a
individual homozygous for the CCR5
deletion allele. Nat Med, 3, 252.
de Roda Husman, A.M., Koot, M., Cornelissen, M., Keet, I.P., Brouwer, M.,
Broersen, S.M., Bakker, M., Roos, M.T., Prins, M., de Wolf, F., Coutinho, R.A.,
Miedema, F., Goudsmit, J., e Schuitemaker, H., 1997, Association between CCR5
genotype and the clinical course of HIV-1 infection. Ann Intern Med, 127(10), 882-890.
Dean, M., Carrington, M., Winkler, C., Huttley, G.A., Smith, M.W.,
Allikmets, R., Goedert, J.J., Buchbinder, S.P., Vittinghoff, E., Gomperts, E.,
Donfield, S., Vlohov, D., Kaslow, R., Saah, A., Rinaldo, C., Detels, R., e
O’Brien, S.J., 1996, Genetics restriction of HIV-1 infection and progression to
AIDS by a deletion allele of the CKR5 structural gene. Science, 273, 1856.
Degerli, N.,
Yilmaz, E., e Bardakci, F., 2005, The Delta32 allele distribution of the CCR5
gene and its relationship with certain cancers in a Turkish population. Clin Biochem, 38(3), 248-252.
D'Souza, M.P., e
Harden, V.A., 1996, Chemokines and HIV-1 second receptors. Confluence of two
fields generates optimism in AIDS research. Nat Med, 2(12), 1293-1300.
Eugen-Olsen, J.,
Iversen, A.K., Garred, P., Koppelhus, U., Pedersen, C., Benfield, T.L.,
Sorensen, A.M., Katzenstein, T., Dickmeiss, E., Gerstoft, J., Skinhoj, P.,
Svejgaard, A., Nielsen, J.O., e Hofmann, B., 1997, Heterozygosity for a
deletion in the CKR-5 gene leads to prolonged AIDS-free survival and slower CD4
T-cell decline in a cohort of HIV-seropositive individuals. AIDS,
11(3),305-310.
Excoffier, L., e Slatkin, M., 1995,
Maximum-likelihood estimation of molecular haplotype frequencies in a diploid
population. Mol Biol Evol, 12(5),921-927.
Galvani, A.P., e
Montgomery, S., 2003, Evaluating plague and smallpox as historical selective
pressure for the CCR5-D32 HIV-resistance allele. PNAS, 100,25.
Guo, S.W., e Thompson, E.A., 1992, Performing the
exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics,
48(2), 361-372.
Gharagozloo, M., Doroudchi, M., Farjadian, S.,
Pezeshki, A.M., Ghaderi, A., 2005, The frequency of CCR5Delta32 and CCR2-64I in
southern Iranian normal population. Immunol Lett, 96(2):277-281.
Harpending,
H.C, e Jenkins, T., 1973, Genetic distance among southern African populations.
In MH Craw ford and PL Workman (eds.): Methods and Theories in anthropologican
Gentetics. Albuquerque University of New Mexico Press; 177 – 199.
Huang, Y., Paxton, W.A., Wolinsky, S.M., Neumann, A.U., Zhang, L., He,
T., Kang, S., Ceradini, D., Jin, Z., Yazdanbakhsh, K., Kunstman, K., Erickson,
D., Dragon, E., Landau, N.R., Phair, J., Ho, D.D., e Koup, R.A., 1996, The role
of a mutant CCR5 allele in HIV-1 trasmission and disease progression. Nat Med,
2, 1240-1243.
Kostrikis, L.G.,
e Martinson, J.J., 1999, Global distribution and origin of the CCR2-64I/CCR5-59653-T
HIV disease-protective haplotype. Conf Retroviruses Opportunistic Infect Jan 31-Feb 4, 6th,174.
Libert, F.,
Cochaux, P., Beckman, G., Samson, M., Aksenova, M., Cao, A., Czeizel, A.,
Claustres, M., de la Rua, C., Ferrari, M., Ferrec, C., Glover, G., Grinde, B.,
Guran, S., Kucinskas, V., Lavinha, J., Mercier, B., Ogur, G., Peltonen, L.,
Rosatelli, C., Schwartz, M., Spitsyn, V., Timar, L., Beckman, L., e Vassart,
G., 1998, The deltaccr5 mutation conferring protection against HIV-1 in
Caucasian populations has a single and recent origin in Northeastern Europe. Hum Mol Genet, 7(3),
399-406.
Liu, R., Paxton, W.A., Choe, S.,
Ceradini, D., Martin, S.R., Horuk, R., MacDonald, M. E., Stuhlmann, H., Koup,
R.A., e Landau, N.R., 1996, Homozygous defect in HIV – 1 coreceptor accounts
for resistance of some multiply – exposed individuals to HIV – 1 infection.
Cell, 86, 367 – 377.
Lucotte, G., e Mercier, G., 1998, Distribution of the
CCR5 gene 32-bp deletion in Europe. J Acquir Immune Defic Syndr Hum Retrovirol, 19(2),
174-177.
Lucotte, G., e Dieterlen, F., 2003, More about the Viking hypotesis of
origin of the D32 mutation in the
CCR5 gene conferring resistance to HIV-1 infection. Infection, Genetics and
Evolution, 3, 293-295.
Martison, J.J.,
Chapman, N.H., Rees, D.C., Liu, Y., e Clegg, J.B., 1997, Global distribution of
the CCR5 gene 32-basepair deletetion. Nature Genetics, 16(1), 100-103.
Martison, J.J.,
Karanicolas, R., Moore, J.P., e Kostrikis, L.G., 2000, Global distribution of
the CCR2-64I/CCR5-59653T HIV-1 disease-protective haplotype. AIDS, 14, 483-489.
Massanet, M.F.,
Castro, J.A., Picornell, A., Misericordia Ramon, M., 1997, Study of the
populations of the Balearic Islands (Spain) using mtDNA RFLPs. Hum Biol, 69(4), 483-498.
Michael, N.L.,
Nelson, J.A., KewalRamani, V.N., Chang, G., O'Brien, S.J., Mascola, J.R.,
Volsky, B., Louder, M., White, G.C. 2nd, Littman, D.R., Swanstrom, R, e
O'Brien, T.R., 1998, Exclusive and persistent use of the entry coreceptor CXCR4
by human immunodeficiency virus type 1 from a subject homozygous for CCR5
delta32. J Virol, 72(7), 6040-6047.
Miguel, A., e
Petitpierre, E., 1989, Red-cell enzyme polymorphisms in Ibiza (Balearic
Islands, Spain). Hum Hered, 39(6),351-355.
Miguel, A.,
Castro, J.A., Ramon, M.M., e Petitpierre, E., 1990, Red cell enzyme
polymorphisms in the Balearic Islands. II. A comparison based on multivariate
analyses. Gene Geogr, 4(2), 89-98.
Morelli, L., Grosso, M.G., Vona, G., Varesi, L., Torroni, A., e
Francalacci, P., 2000, Frequency distribution of mitochondrial DNA haplogroups
in Corsica and Sardinia. Hum Biol, 72(4):585-595.
Moral, P. 1986, Estudio antropogenetico de diversos polimorpfismos
hematològicos en la isla de Menorca. Ph D. Thesis, Universitat de Barcelona, Barcelona,
Spain.
O'Brien, T.R., Winkler, C., Dean, M., Nelson, J.A., Carrington, M.,
Michael, N.L., e White, G.C. 2nd., 1997, HIV-1 infection in a man homozygous
for CCR5 delta 32. Lancet, 349(9060), 1219.
Ohta, T., e Kimura, M., 1969, Linkage disequilibrium at steady state determined by random genetic
drift and recurrent mutation. Genetics, 63, 229 – 238.
Piazza, A.,
Cappello, N., Olivetti, E., e Rendine S., 1988, A genetic history of Italy. Ann Hum Genet, 52 (3):203-213.
Picornell, A.,
Miguel, A., Castro, J.A., Misericordia Ramon, M., Arya, R., e Crawford, M.H.,
1996, Genetic variation in the population of Ibiza (Spain): genetic structure,
geography, and language. Hum Biol, 68(6), 899-913.
Rana, S.,
Besson, G., Cook, D.G., Rucker, J., Smyth, R.J., Yi, Y., Turner, J.D., Guo,
H.H., Du, J.G., Peiper, S.C., Lavi, E., Samson, M., Libert, F., Liesnard, C.,
Vassart, G., Doms, R.W., Parmentier, M., e Collman, R.G., 1997, Role of CCR5 in
infection of primary macrophages and lymphocytes by macrophage-tropic strains
of human immunodeficiency virus: resistance to patient-derived and prototype
isolates resulting from the delta ccr5 mutation. J Virol, 71(4),3219-3227.
Rappaport, J.,
Cho, Y.Y., Hendel, H., Schwartz, E.J., Schachter, F., e Zagury, J.F., 1997, 32
bp CCR-5 gene deletion and resistance to fast progression in HIV-1 infected
heterozygotes. Lancet, 349(9056), 922-923.
Raymond, M., e Rousset, F., 1995, Genepop (ver. 1.2):
a population genetics software for exact tests and ecumenicism. J Hered, 86,
248-249.
Samson, L.,
Libert, F., Doranz, B.J., Rucker, J., Liesnard, C., Farber, C.M., Saragosti,
S., Lapoumeroulie, C., Cognaux, J., Forceille, C., Muyldermans, G.,
Verhofstede, C., Burtronboy, G., George, M., Imai, T., Rana, S., Yi, Y., Smyth,
R.J., Colliman, R.G., Doms, R.W., Vassart, G., e Parmentier, M., 1996,
Resistance to HIV-1 infection in Caucasian individuals bearing mutant alleles
of the CCR-5 chemokine receptor gene. Nature, 382, 722.
Schneider, S.,
Roessli, D., e Excoffier, L., 2000, Arlequin: a software for population
genetics data analysis (Ver 2.000). Genetics and Biometry Lab, Dept. of
Anthropology, University of Geneva.
Scozzari, R.,
Cruciani, F., Pangrazio, A., Santolamazza, P., Vona, G., Moral, P., Latini, V.,
Varesi, L., Memmi, M., Romano, V., De Leo, G., Gennarelli, M., Jaruzelska, J.,
Villems, R., Parik, J., Macaulay, V., e Torroni, A., 2001, Human Y-chromosome
variation in the western Mediterranean area: implications for the peopling of the region. Hum Immunol, 62(9):871-884.
Smith, M.W., Dean, M., Carrington, M., Winkler, C., Huttley, G.A., Lomb,
D.A., Goedert, J.J., O'Brien, T.R., Jacobson, L.P., Kaslow, R., Buchbinder, S.,
Vittinghoff, E., Vlahov, D., Hoots, K., Hilgartner, M.W., e O'Brien, S.J.,
1997, Contrasting genetic influence of CCR2 and CCR5 variants on HIV-1
infection and disease progression. Science, 1997, 277(5328), 959-965.
Stephens, J.C., Reich, D.E., Goldstein, D.B., Shin,
H.D., Smith, M.W., Carrington, M., Winkler, C., Huttley, G.A., Allikmets, R.,
Schriml, L., Gerrard, B., Malasky, M., Ramos, M.D., Morlot, S., Tzetis, M.,
Oddoux, C., Di Giovine, F. S., Nasioulas, G., Chandler, D., Aseev, M., Hanson,
M., Kalaydjieva, L., Glavac, D., Gasparini, P., Kanavakis, E., Claustres, M.,
Kambouris, M., Ostrer, H., Duff, G., Baranov, V., Sibul, H., Metspalu, A.,
Goldman, D., Martin, N., Duffy, D., Schmidtke, J., Estivill, X., O’Brien, S. J.,
e Dean, M., 1998, Dating the Origin of the CCR5-D32 aids-Resistance
Allele by the Coalescence of Haplotypes. Am J Hum Genet, 62, 1507-1515.
Struyf, F.,
Thoelen, I., Charlier, N., Keyaerts, E., Van der Donck, I., Wuu, J., e Van
Ranst, M., 2000, Prevalence of CCR5 and CCR2 hiv-coreceptor gene polymorphisms
in Belgium. Hum Hered, 50, 304-307.
Su, B., Jin, L.,
Hu, F., Xiao, J., Luo, J., Lu, D., Zhang, W., Chu, J., Du, R., Geng, Z., Qiu,
X., Xue, J., Tan, J., O'Brien, S. J., e Chakraborty, R., 1999, Distribution of
Two HIV-1-Resistance Polymorphism (SDF1-3’A and CCR2-64 I) in East Asian and
population and Its Implication in AIDS Epidemiology. Am J Hum Genet, 65, 1047-1053.
Su, B., Sun, G.,
Lu, D., Xiao, J., Hu, F., Chakraborty, R., Deka, R., e Jin, L., 2000, Distribution
of three HIV-1 resistance-conferring polymorphism (SDF1-3’A, CCR2-64 I, and
CCR5-D32) in global population. Eur J Hum Genet, 8, 975-979.
Theodorou, I., Meyer, L., Magierowska, M., Katlama, C., e Rouzioux, C.,
1997, HIV-1 infection in an individual homozygous for CCR5 delta 32. Lancet, 349(9060), 1219-1220.
Torroni, A., Bandelt, H.J., D'Urbano, L.,
Lahermo, P., Moral, P., Sellitto, D., Rengo, C., Forster, P., Savontaus, M.L.,
Bonne-Tamir, B., e Scozzari, R., 1998, mtDNA analysis reveals a major late Paleolithic
population expansion from southwestern to northeastern Europe. Am J Hum Genet, 62(5):1137-1152.
Tofanelli, S.,
Taglioli, L., Varesi, L., e Paoli, G., 2004, Genetic history of the population
of Corsica (western Mediterranean) as inferred from autosomal STR analysis. Hum Biol, 76(2):229-251.
Varesi, L., Memmi, M., Cristofari, M.C., Mameli, G.E., Calò, C.M., e Vona,
G., 2000, Mitochondrial control-region sequence variation in the Corsican
population, France. Am J Hum Biol, 12(3), 339-351.
Vona, G., Salis,
M., Bitti, P., e Succa, V., 1994, Blood groups of the sardinian population
(Italy). Anthrop Anz, 52, 4, 297-304.
Vona, G., 1995, Le caratteristiche genetiche dei sardi. Antropologia
contemporanea. 18 (3-4), 101 –109.
Vona, G., 1997,
The peopling of Sardinia (Italy): History and effects. Int J Anthropol, 12, 71 – 87.
Vona, G., Memmì,
M., Calò, C.M., Latini, V., Vacca, L., Succa, V., Ghiani, M.E., Moral, P., e
Varesi, L., 2002, Genetic structure of the Corsican population (France) : a
rewiew. Recent Res Devel Human Genet, 1, 147-164.
Vona, G., Moral,
P., Memmi, M., Ghiani, M.E., e Varesi, L., 2003, Genetic structure and
affinities of the Corsican population (France): classical genetic markers
analysis. Am
J Hum Biol, 15(2), 151-163.