
Peña, J.A., Alfonso-Sánchez, M.A., Pérez-Miranda, A.,
García-Obregón, S., 2004, Flujo génico y presiones selectivas en Europa. Una
aproximación a partir de clinas de ADN codificante y no codificante. Antropo,
8, 117-122. www.didac.ehu.es/antropo
Flujo
génico y presiones selectivas en Europa. Una aproximación a partir de clinas de
ADN codificante y no codificante
Gene flow and selective pressures in Europe. An approach from genic and nongenic DNA clines
Peña,
J.A., Alfonso-Sánchez, M.A., Pérez-Miranda, A., García-Obregón, S.
Dpto.
Genética, Antropología Física y Fisiología Animal. Universidad del País Vasco. Aptdo. 644. 48080 Bilbao. Spain. E-mail:
ggppegaj@lg.ehu.es
Palabras clave: clinas, flujo génico, deriva genética, presiones
selectivas.
Key-words: clines, gene flow, genetic drift, selective pressures.
Resumen
En este trabajo se aborda un análisis
filogeográfico de las poblaciones europeas a partir de una base de datos
compuesta por frecuencias de polimorfismos de ADN codificante y no codificante.
Entre los primeros se incluyen grupos sanguíneos, proteínas, haplotipos GM y
loci HLA. Entre los no codificantes se consideran secuencias de ADN repetitivo
de minisatélites y microsatélites.
Tanto la detección de las clinas como la determinación de su
orientación se han realizado mediante sucesivos análisis de correlación entre
las frecuencias alélicas o haplotípicas y la posición geográfica de las
poblaciones en un sistema de coordenadas móviles, donde la latitud y la
longitud han rotado entre 0 y 360 grados.
No se han detectado diferencias en la distribución espacial
de las clinas entre marcadores codificantes y no codificantes.
En la discusión de los patrones de distribución geográfica
de las frecuencias génicas se han considerado diferentes presiones evolutivas: i) el flujo génico en el
poblamiento de Europa, considerando tanto la hipótesis de la recolonización
Postglacial desde los refugios del sur del continente como la hipótesis de la
difusión démica de los granjeros del Neolítico desde el Próximo Oriente, y ii) la existencia de presiones
selectivas que puedan haber generado un cierto grado de microdiferenciación
genética
Abstract
In
this study, a phylogeographic analysis for the European context is carried out
using a database with frequencies for both, genic and nongenic fragments of
DNA. Among the genic fragments of DNA, serological markers including blood
groups, proteins, GM and HLA loci were considered. Among the second ones,
repetitive DNA sequences from minisatellites (VNTR) and microsatellite (STR)
loci were considered.
The
direction of allelic frequency clines detected for these genomic regions was
determined by analyzing the correlations between gene frequencies and the
geographic position of the populations, in a system of moving coordinates where
the latitude and the longitude rotated between 0 and 360 degrees.
No
differences in the spatial distribution of clines between genic and nongenic
markers were found.
Various
evolutionary forces were considered in discussing the origin of the spatial
structuring of the allele frequencies: i) gene flow,
argued from the hypotheses of Post-glacial recolonization from southern Europe
or the demic diffusion of farmers from the Near East into Europe; and ii) the
existence of selective pressures that could have generated genetic
microdifferentiation.
Introduction
El origen de una clina puede deberse a una o varias de las
siguientes causas:
i) la existencia de dos
poblaciones con diferentes origenes ubicadas en los extremos de una amplia
región geográfica,
ii) un proceso combinado de
flujo génico y deriva, y
iii) una presión selectiva de
incidencia variable a lo largo de una dimensión espacial.
En este trabajo se ha analizado la orientación de las clinas
detectadas en dos grupos de marcadores, uno compuesto por genes y otro por
secuencias de ADN repetitivo.
El objetivo es identificar la posible existencia de un
efecto fundador asociado a un proceso de flujo génico en el contexto del debate
acerca
del origen del patrimonio genético de las poblaciones europeas.
Materiales y métodos
Se ha recopilado una base de datos con más de 2500
frecuencias de los genes ABO, ACP1, ADA, AK1, Duffy, GC, GM, HLA*A, HLA*B, Hp,
Kidd, MNSs, P1 , PI, Rh, y Tf, y otra base de datos con más de 3800 frecuencias
de los loci repetitivos APOB, D1S80, D3S1358, D5S817, D7S820, D8S1179, D13S317,
D17S5, FGA, THO01, and VW31A.
Con el fin de minimizar las desviaciones debidas a errores
de muestreo, se han considerado regiones geográficas de similar tamaño. Dichas
regiones pueden incluir un país entero cuando su tamaño es pequeño, como en el
caso de Holanda o Bélgica, o incluir sólo una parte cuando el país es grande.
En todos los casos, cuando se ha contado con varias muestras de una misma area,
se han calculado sus correspondientes frecuencias alélicas como el valor medio
de todas las frecuencias disponibles. Se han excluido aquellos trabajos en los
que el tamaño muestral fuese muy reducido.
Para la identificación de las posibles
clinas se ha calculado el coeficiente de correlación de Pearson entre las
frecuencias génicas y las coordenadas geográficas de las diferentes poblaciones
en un sistema de coordenadas móviles en el que la latitud y la longitud han
rotado de 0 a 360 grados. De este modo, cuando existe un gradiente clinal, se
detecta la orientación y el sentido en los que la significación de la
correlación es máxima.
La comparación de la distribución de las orientaciones de las clinas entre loci codificantes y no codificantes se ha realizado mediante el test no paramétrico de Mann-Whitney. Después, la posible existencia de un origen común de ambas distribuciones se evaluó mediante el coeficiente de correlación no paramétrico de Spearman.
Resultados
Se han detectado 45 clinas en alelos correspondientes
a genes y 49 en alelos de secuencias repetitivas. La distribución de
frecuencias de la orientación de las clinas se muestra en las figuras 1 y 2.
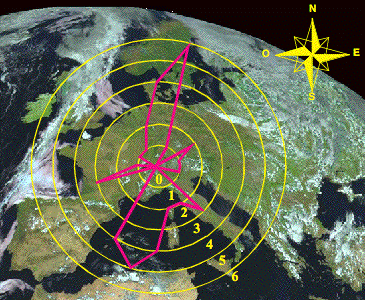
Figura 1. Gráfico
direccional con el número de clinas y su orientación en Europa para un grupo de
genes codificantes, incluyendo grupos sanguíneos, proteínas séricas, haplotipos
GM y loci HLA.
Figure 1. Directional
plot showing the pattern of gradients for a group of blood groups, proteins, GM
haplotypes and HLA loci.
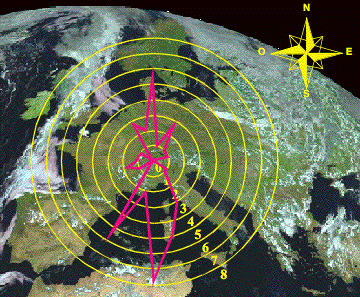
Figura 2. Gráfico
direccional con el número de clinas y su orientación en Europa para un grupo de
loci de ADN repetitivo.
Figure 2. Directional
plot showing the pattern of gradients for a group of VNTR and STR loci.
Tanto en el grupo de loci codificantes
como entre los no codificantes se observa un predominio de clinas
latitudinales, que superan con mucho el número de clinas longitudinales. De
este modo, la mayor parte de los gradientes significativos detectados muestran
una tendencia creciente (o decreciente, en su caso) en sentido Norte o
Nor-nordeste.
El test de Mann-Whitney no mostró
diferencias significativas entre ambas distribuciones (U: 872; z: -1,746; p:
n.s.), de modo que puede asumirse que son similares. Por su parte, el
coeficiente de correlación de Spearman mostró una asociación significativa (![]() :0,562; g.l: 23; p<0,01), por lo que cabría esperar una
causa común, no fruto del azar, en la distribución de las clinas en ambos
grupos de polimorfismos.
:0,562; g.l: 23; p<0,01), por lo que cabría esperar una
causa común, no fruto del azar, en la distribución de las clinas en ambos
grupos de polimorfismos.
Discusión
Se han descrito dos hipótesis
alternativas basadas en mecanismos no adaptativos para explicar el patrón de
distribución de las frecuencias génicas en las poblaciones europeas actuales.
La primera en ser propuesta fue la
hipótesis de la difusión démica del Neolítico (Menozzi et al, 1978; Ammerman y
Cavalli-Sforza, 1984). Los granjeros provenientes de uno de los más importantes
focos de origen y expansión del Neolítico, el creciente fértil en el Próximo
Oriente, portadores de una ventaja cultural muy importante sobre los hombres
del Paleolítico, habrían colonizado Europa desplazando o absorbiendo a
aquellos, que quedarían diluídos en un patrimonio genético mucho más grande,
fruto de la mayor densidad de población que pueden alcanzar las poblaciones
agricultoras y ganaderas en relación a las cazadoras-recolectoras (figura 3).
En consecuencia, la tendencia predominante en el patrimonio genético de las
poblaciones humanas del continente europeo, consecuencia de este flujo
migratorio, debiera verse reflejada en la forma de clinas longitudinales. Entre
otros trabajos que apoyan esta hipótesis pueden citarse los de Semino et al. (1996), Cavalli-Sforza y
Minch (1997), Chikhi et al. (1998), Quintana-Murci et al. (1999) y Simoni et al. (2000).
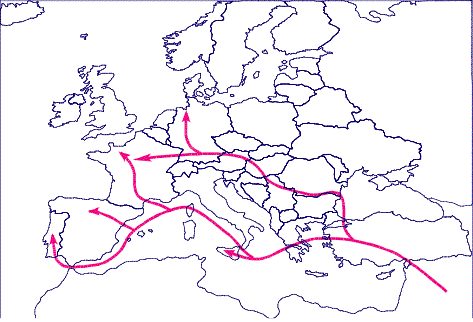
Figura 3. La difusión
démica del los granjeros del Neolítico en su dispersión desde el Próximo
Oriente.
Figure 3. The demic diffusion of Neolithic farmers dispersing from the Near East.
La hipótesis de la recolonización
postglacial del Paleolítico, por el contrario, minimiza el efecto que la
llegada del Neolítico a Europa pudo tener sobre el patrimonio genético de sus
habitantes. En este caso se considera que el proceso más influyente pudo ser la
migración desde los refugios del sur de los últimos cazadores-recolectores de
finales del Paleolítico, ocupando nuevamente todo el continente tras el
retroceso de los hielos al final de la última glaciación (Richards
et al., 1996;
Torroni et al.,
1998; Richards, 2003). El aumento de la densidad de sus pobladores se daría
gradualmente conforme éstos adquiriesen las prácticas agrícolas provenientes
del Este. Así, las clinas que pudieran observarse, de acuerdo a esta hipótesis,
debieran ser fundamentalmente latitudinales.
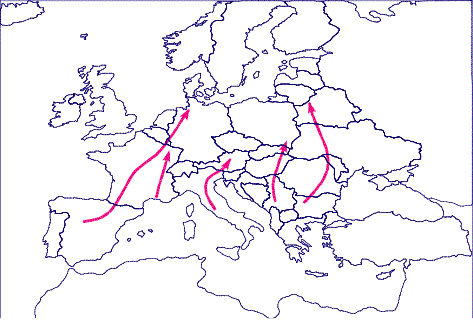
Figura
4. Flujo génico desde los refugios del sur de Europa
tras el final de la última glaciación.
Figure
4. The gene flow across the continent from the southern
European refugium after the Last Glacial Maximum
A la luz de los resultados obtenidos
en este trabajo, habiéndose detectado la existencia de una mayoría de clinas
latitudinales tanto en genes codificantes como en minisatélites y
microsatélites, se sugiere un proceso de flujo génico y deriva con esa misma
orientación, lo que concuerda con las migraciones del Paleolítico que
probablemente ocurrieron después de la última glaciación.
No obstante, no puede descartarse la
existencia de presiones selectivas de intensidad variable en sentido
latitudinal, que puedan quedar ocultas por corresponderse con la misma
orientación con el conjunto de gradientes descritos, como ha sido revelado para
la persistencia de actividad lactasa (Peña et al, 2002).
Agradecimientos. Este
trabajo ha sido financiado por el proyecto UPV 00154.310-E-14873/2002 de la
Universidad del País Vasco. M.
Alfonso-Sánchez ha sido financiado por una beca postdoctoral del "Programa
de Formación de Investigadores" del Departamento de Educación,
Universidades e Investigación del Gobierno Vasco.
Bibliografía
Ammerman, A. J.,
Cavalli-Sforza, L. L., 1984, The Neolithic Transition and the Genetics of
Population in Europe. (Princeton, USA: Princeton University Press).
Cavalli-Sforza, L. L., Minch,
E., 1997, Paleolithic and Neolithic lineages in the European mitochondrial gene
pool, Am. J. Hum. Genet., 61, 247-251.
Chikhi, L., Destro-Bisol, G., Bertorelle, G., Pascali, V., Barbujani, G., 1998, Clines of nuclear DNA markers suggest a largely Neolithic ancestry of the European gene pool. Proc. Natl. Acad. Sci. USA, 95, 9053-9058.
Menozzi, P., Piazza, A.,
Cavalli-Sforza, L. L., 1978, Synthetic maps of human gene frequencies in
Europeans. Science, 201, 786-792.
Peña, J. A., Alfonso-Sánchez,
M. A., García-Obregón, S., Pérez-Miranda, A. M., 2002,
Persistencia de actividad lactasa en población residente en el País
Vasco. Antropo, 3, 51-60. www.didac.ehu.es/antropo
Quintana-Murci, L., Semino,
O., Minch, E., Passarimo, G., Brega, A., Santachiara-Benerecetti, A. S., 1999,
Further characteristics of proto-European Y chromosomes, European Journal of
Human Genetics, 7, 603-608.
Richards, M., 2003. The
Neolithic invasion of Europe. Annu. Rev. Anthropol. 32: 135-162.
Richards, M., Côrte-Real, H.,
Forster, P., Macaulay, V., Wilkinson-Herbots, H., Demaine, A., Papiha, S., et
al (1996) Paleolithic and Neolithic lineages in the European mitochondrial gene
pool. Am. J. Hum. Genet., 59, 185-203.
Semino, O., Passarino, G.,
Brega, A., Fellous, M., Santachiara-Benerecetti, A. S., 1996, A view of the
Neolithic demic diffusion in Europe through two Y chromosome-specific markers.
Am. J. Hum. Genet., 59, 964-968.
Simoni, L., Calafell, F., Pettener, D., Bertranpetit, J.,
Barbujani, G., 2000, Geographic patterns of mtDNA diversity in Europe. Am. J.
Hum. Genet., 66, 262-278.
Torroni, A., Bandelt, H., Dúrbano, L., Lahermo, P., Moral, P., Sellito, D., Rengo, C., Forster, P., Savontaus, M., Bonné-Tamir, B., Scozzari, R., 1998, mtDNA analysis reveals a major Late Paleolithic population expansion from Southwestern to Northeastern Europe. Am. J. Hum. Genet., 62, 1137-1152.