
Colloques du Groupement des Anthropologistes de Langue Française (GALF)
Santovito, A., Marin, A., Zampicinini, G.P., Girotti, M., Bigatti, M.P., Lamberti, L., Del Pero, M., Selvaggi, A., Cervella, P., 2004, Polymorphismes par insertion Alu dans trois populations du Nord de l’Italie. Antropo, 7, 45-53. www.didac.ehu.es/antropo
Polymorphismes par insertion Alu dans trois populations du Nord de l’Italie
Alu polymorphisms in three populations from Northern Italy
Santovito, A., Marin, A., Zampicinini, G.P., Girotti, M., Bigatti, M.P., Lamberti, L., Del Pero, M., Selvaggi, A., Cervella, P.
Département de Biologie Animale et de l’Homme, Université de Turin, Via Accademia Albertina 17, 10123 – Turin (Italie)
Corrispondenza: Alfredo Santovito. Département de Biologie Animale et de l’Homme, Université de Turin, Via Accademia Albertina 17, 10123 – Turin (Italie). E-mail: alfredo.santovito@unito.it
Mots-clés: Alu, polymorphismes, marqueurs génétiques, PCR.
Keywords: Alu, polymorphisms, genetic markers, PCR.
Résumé
Le polymorphisme à six différents locus d'insertion de séquences Alu, (ACE, APO, FXIIIB, TPA25, A25 et B65) a été examiné dans trois populations humaines du Piémont nord-oriental: Postua, un village de 570 habitants à isolement géographique et culturel partiel, fermé à l’immigration et avec un degré élevé de mariages entre consanguins, et, comme contrôle, deux agglomérations urbaines limitrophes, Cavaglià, un pays agricole de 3.680 habitants et Biella, une ville industrielle de 47.000 habitants.
Des déviations significatives de l’équilibre Hardy-Weinberg n'ont été observées, à l'exception du locus ACE dans les populations de Postua et de Cavaglià et, pour le locus B65, dans celle de Biella. Dans 5 des 6 locus analysés, Postua montrait les valeurs de fréquence d'insertion les plus basses par rapport aux deux autres populations analysées. De plus, Postua montrait une valeur d'hétérozygotie moyenne plus basse (0.291±0.003) que celles de Cavaglià (0.329±0.002) et de Biella (0.343±0.003). La valeur moyenne de FST était basse (0,038), indiquant par là que la variance des valeurs de fréquence d'insertion Alu est essentiellement due à des différences à l’intérieur des populations et non pas entre les populations.
Les données de fréquence d'insertion Alu ont été étudiées conjointement avec celles que l'on trouve en littérature pour 7 autres populations européennes et l'on en a tiré un arbre Neighbor-Joining où Postua a une position basale, suivie de Cavaglià et Biella. De même, l’analyse de la composante principale situe Postua dans une position relativement isolée par rapport à Cavaglià et à Biella ainsi que par rapport aux autres populations européennes.
Il en résulte, pour Postua, une population à basse hétérogénéité génétique, caractérisée, dans presque tous les locus analysés, par une fréquence d'insertions Alu et par des valeurs d'hétérozygotie moyenne plus basses que celles des populations utilisées comme contrôle. Des facteurs culturels, tels que l'habitude de contracter mariage entre membres de la même communauté avec, en conséquence, une augmentation de la consanguinité entre les habitants et un certain isolement que Postua a maintenu - dans le passé du moins – pourraient avoir contribué à déterminer la situation actuelle de cette population.
Abstract
Polymorphisms at six distinct Alu insertion loci (ACE, APO, FXIIIB, TPA25, A25 and B65) were examined in three north-east Piedmont human populations: Postua, a village of 570 inhabitants with partial geographical and, in the past, cultural isolation, closed to immigration, with a high degree of marriages between relatives; as control, two neighbouring urban agglomerates, Cavaglià, an agricultural town of 3,680 residents, and Biella, an industrial city of 47,000 inhabitants, were considered.
Significant deviations from the Hardy-Weinberg equilibrium were never observed, except for the locus ACE in Postua and Cavaglià populations and for the locus B65 in Biella. Five loci out of six showed for Postua insertion frequency values lower than those observed for Cavaglià and Biella. Postua presented, for all loci, an average heterozygosity (0.291±0.003) lower than Cavaglià (0.329±0.002) and Biella (0.343±0.003). The average FST value were low (0,038), indicating that the variance of Alu insertion frequencies is mainly due to differences within populations rather than between populations.
The Alu insertion frequency data were analysed along with data available in the literature relatively to other 7 European populations, and a Neighbor-Joining tree was constructed: Postua occupied a basal position, followed by Cavaglià and Biella. Moreover, the principal components analysis places Postua in a relatively isolated position with respect to Cavaglià, Biella and the other European populations.
Our analysis depicts Postua as characterised by low genetic heterogeneity, with lower Alu insertion frequencies and average heterozigosity values, compared to the control populations. Cultural factors, like the tradition to get married within the community, with consequent increasing of the consanguinity among inhabitants, and a partial isolation that Postua maintained, at least in the past, could have contributed to determine the current genetic structure of this population.
Introduction
Les éléments Alu sont une famille de courtes séquences d'ADN répétitif, dispersées à l’intérieur du génome des primates et dérivant d'une rétrotransposition de l’ARN 7SL (Ullu et al., 1982), un abondant ARN cytoplasmique, transcrit par l'ARN-polymérase III, et qui est un composant essentiel de la particule d'identification du signal (SRP) des eucaryotes et permet le transport des protéines de sécrétion à travers le réticulum endoplasmique. De même que tous les ARN transcrits par l'ARN-polymérase III, l'ARN 7SL contient, en son sein, le promoteur de la transcription. Cela a rendu possible la transcription et l’ultérieure rétrotransposition de copies d'éléments Alu insérés dans le génome.
Les éléments Alu sont présents en environ 1.200.000 copies à l'intérieur du génome humain et chaque répétition Alu est longue d'environ 300 nucléotides (Weiner et al., 1986; Okada, 1994). L’amplification d'éléments Alu dans le génome humain est un processus encore en cours, comme le prouvent les pathologies causées par l'insertion d'éléments Alu à l'intérieur de gènes spécifiques (Deininger et Baitzer, 1999). Les éléments Alu localisés à l'intérieur du génome ne sont généralement pas sujets à des pertes ou à des réarrangements, et ils représentent donc des marqueurs génétiques stables du chromosome lui-même.
Le polymorphisme d'insertion Alu consiste dans la présence/absence d'un élément Alu dans un locus chromosomique particulier, phénomène pouvant être aisément mis en évidence par PCR (Batzer et al., 1994; Hammer, 1994; Kass et al., 1994). Ce polymorphisme d'insertion est particulièrement utile dans les études de génétique de population, car la probabilité de rétrotransposition indépendante, dans le même site chromosomique, est virtuellement nulle (Batzer et Deininger, 1991). Par conséquent, tous les locus portant une insertion Alu polymorphique particulière dérivent d'un seul événement et sont donc identiques par descendance (Batzer et al, 1994).
Pour les insertions Alu, l'état primitif Alu est l'absence de l'insertion, tandis que l’insertion de l’élément Alu dans un locus déterminé représente le changement mutationnel. Cette connaissance de l'état ancestral et de la direction du changement mutationnel facilite grandement l’analyse des relations entre populations, ce qui est généralement plus difficile si l'on utilise d'autres types de marqueurs (par ex., RFLP, VNTR et microsatellites).
Dans le cadre d'un projet d'étude sur des populations de la chaîne des Alpes occidentales, nous avons analysé 6 locus (ACE, APO, FXIIIB, TPA25, A25 et B65) intéressés par des phénomènes d'insertion Alu. Nous avons considéré 3 populations limitrophes situées dans une région pré-alpine du Piémont oriental: Postua, un petit pays de 570 habitants à peine, caractérisé par un isolement partiel dû à des causes aussi bien géographiques que culturelles; Biella, ville industrielle de 47.000 habitants, et Cavaglià, pays agricole de 3.680 habitants, situé dans une position géographique intermédiaire entre Postua et Biella.
La position géographique de Postua, éloignée des voies de passage les plus fréquentées de la zone, a favorisé un certain isolement de cette communauté, laquelle a vécu - et en partie vit encore - renfermée en elle-même, dans la perpétuation d'un mode de vie fondé sur les traditions locales et les activités agricoles sédentaires. De plus, bien que les habitants de Postua aient participé à des vagues migratoires (vers la France surtout), les émigrés et leurs descendants sont restés en contact avec leur pays d'origine: jusqu'à la deuxième guerre mondiale, les émigrés retournaient presque toujours à Postua pour se marier. Il est donc possible de parler de “familles de Postua” dans le cas, aussi, des noyaux familiaux des émigrés. Cela rend la population de Postua relativement “fermée” du point de vue du flux génique et donc intéressante pour des études de population.
Le but de notre travail a été de vérifier, sur la base des polymorphismes d'insertion de séquences Alu, l'existence d'une trace génétique de cet isolement.
Matériels et méthodes
Populations étudiées
L'échantillon que nous avons analysé était constitué de 250 individus: 102 originaires de Postua, 95 de Biella et 53 de Cavaglià.
Le groupe de Postua était constitué de sujets indigènes depuis au moins 3 générations, tandis que pour Biella et Cavaglià ont été utilisés des échantillons provenant de donneurs de sang. Pour autant que nous le sachions, les individus examinés n'étaient pas parents, à l'exception de la population de Postua pour laquelle a été observée la répétition de certains noms de famille parmi les personnes qui se sont volontairement soumises à la prise de sang, et pour lesquelles est, en tout cas, inconnu le degré de parenté.
Analyse biodémographique
Nous avons examiné les registres matrimoniaux de Postua de 1866 à 1995, pour un total de 1.089 actes de mariage. Les données collectées ont été introduites dans une base de données, en utilisant le programme WinFamy (De Iasio, 1999), et elles ont été ensuite traitées pour calculer les valeurs de l'isonymie observée, de l'isonymie escomptée et du coefficient de consanguinité.
Extraction et amplification des échantillons
Les échantillons de sang (5-10 ml de sang périphérique)
ont été recueillis dans des éprouvettes stériles contenant de l'héparine. L'ADN
a été obtenu à partir de 3 ![]() l selon un protocole utilisant une suspension à 5% de la
résine à échange ionique Chelex (Walsh P.Ss et al., 1991). Chaque échantillon
d'ADN a été ensuite amplifié par PCR en utilisant des primer locus spécifiques
(Tableau 1). La réaction de PCR a été effectuée dans un volume total de 25
l selon un protocole utilisant une suspension à 5% de la
résine à échange ionique Chelex (Walsh P.Ss et al., 1991). Chaque échantillon
d'ADN a été ensuite amplifié par PCR en utilisant des primer locus spécifiques
(Tableau 1). La réaction de PCR a été effectuée dans un volume total de 25 ![]() l, contenant 10
l, contenant 10 ![]() l d'ADN, conformément aux conditions indiquées en littérature
pour les différents locus (Arcot et al., 1995). Les produits d'amplification
ont été ensuite analysés et visualisés sur gel d'agarose à 2%.
l d'ADN, conformément aux conditions indiquées en littérature
pour les différents locus (Arcot et al., 1995). Les produits d'amplification
ont été ensuite analysés et visualisés sur gel d'agarose à 2%.
|
Alu insertion |
5’ Primer sequence (5’-3’) |
3’ Primer sequence (5’-3’) |
Ta(°C) |
|
TPA25 |
GTAAGAGTTCCGTAACAGGACAGCT |
CCCCACCCTAGGAGAACTTCTCTTT |
58 |
|
ACE |
CTGGAGACCACTCCCATCCTTTCT |
GATGTGGCCATCACATTCGTCAGAT |
58 |
|
APO |
AAGTGCTGTAGGCCATTTAGATTAG |
AGTCTTCGATGACAGCGTATACAGA |
50 |
|
FXIIIB |
TCAACTCCATGAGATTTTCAGAAGT |
CTGGAAAAAATGTATTCAGGTGAGT |
56 |
|
B65 |
ATATCCTAAAAGGGACACCA |
AAAATTTATGGCATGCGTAT |
52 |
|
A25 |
CCACAAATAGGCTCATGTAGAAC |
TATATTATGGCCTGGATTATACC |
57 |
Tableau 1. Alu, localisation chromosomique, séquences et températures d’utilisation des primers
Table 1. Alu, chromosomal location, primer sequence and annealing temperatures
Analyse statistique
Pour le calcul des fréquences alléliques, de leur accord avec l'équilibre Hardy Weinberg et de la valeur de FST, pour chaque couple de population a été utilisé le programme GENEPOP (Guo et al., 1992). Par la suite, le programme NEIGHBOR faisant partie du paquet PHYLIP 3.5c (Felsensein, 1989) a été utilisé pour construire un arbre représentant les relations entre les populations. Les relations entre les populations ont été ultérieurement examinées à travers l’analyse de la composante principale (PC) effectuée en utilisant le programme SYSTAT 1.0 (SPSS Inc. 1996).
Résultats
Dans la figure 1 est représentée l'évolution de l'isonymie observée et de l'isonymie escomptée: comme on le voit, l'isonymie matrimoniale observée présente un pic dans la première décennie examinée, pour diminuer ensuite constamment. Puis l'on remarque une deuxième augmentation se rapportant à la période 1926-1935. A partir de 1950, l'on ne constate plus de cas de mariages isonymiques.
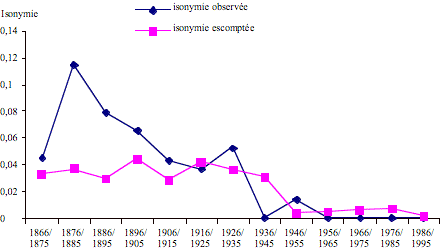
Figure 1. Population de Postua: isonymie observée et escomptée pendant la période 1866 – 1995.
Figure 1. Postua population: observed and theoretical isonomy during the period 1866-1995.
Le coefficient de consanguinité (figure 2) présente une évolution extrêmement semblable à celle de l'isonymie matrimoniale observée. Dans ce cas aussi, nous trouvons un pic initial d'augmentation, suivi d'une diminution progressive, toutefois interrompue par une augmentation momentanée au cours de la décennie 1926/35.
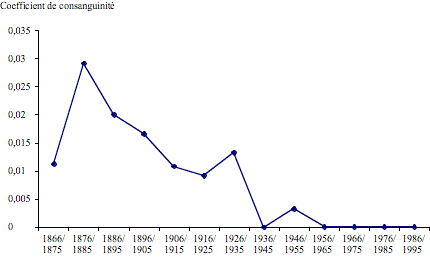
Figure 2. Population de Postua: coefficient de consanguinité au cours de la période 1866 - 1995.
Figure 2. Postua population: coefficient of consanguinity during the period 1866-1995.
Population |
n |
ACE |
APO |
FXIIIB |
TPA25 |
A25 |
B65 |
Postua |
102 |
0.338 |
0.951 |
0.422 |
0.353 |
0 |
0.157 |
|
Cavaglià |
53 |
0.434 |
0.953 |
0.396 |
0.575 |
0.075 |
0.198 |
|
Biella |
95 |
0.537 |
0.989 |
0.453 |
0.595 |
0.058 |
0.347 |
Tableau 2. Fréquences d'insertion Alu dans les populations de Postua, Cavaglià et Biella (n = nombre d'individus)
Table 2. Alu insertion frequencies in Postua, Cavaglià and Biella populations
Dans le tableau 2 sont représentées les données relatives aux fréquences d'insertion Alu des trois populations étudiées: l'on observe que dans 5 (ACE, APO, TPA25, A25 et B65) des 6 locus c'est Postua qui présente les valeurs les plus basses.
Cette différence est particulièrement évidente pour les locus ACE et TPA25.
Les fréquences alléliques sont conformes à
l’équilibre Hardy-Weinberg, à l'exception du locus ACE pour Postua et Cavaglià
(![]() = 6.594, 0.02<p<0.05 pour Postua et
= 6.594, 0.02<p<0.05 pour Postua et ![]() = 27,245
p<0.001 pour Cavaglià), et du locus B65 pour Biella (
= 27,245
p<0.001 pour Cavaglià), et du locus B65 pour Biella (![]() = 17.9 p<0.001). Ces déviations représentent probablement
des fluctuations statistiques occasionnelles.
= 17.9 p<0.001). Ces déviations représentent probablement
des fluctuations statistiques occasionnelles.
|
Population |
n |
ACE |
APO |
FXIIIB |
TPA25 |
A25 |
B65 |
Hétm±ES |
|
Postua |
102 |
0,448 |
0,093 |
0,488 |
0,457 |
0 |
0,265 |
0,291±0.003 |
|
Cavaglià |
53 |
0,496 |
0,055 |
0,478 |
0,489 |
0,140 |
0,318 |
0,329±0.002 |
|
Biella |
95 |
0,497 |
0,021 |
0,496 |
0,482 |
0,109 |
0,453 |
0,343±0.003 |
|
FST/locus |
|
0,044 |
0,009 |
0,004 |
0,075 |
0,037 |
0,059 |
|
Tableau 3. Valeurs d’hétérozygotie dans les populations de Postua, Cavaglià et Biella (n = nombre d'individus ; Hétm = Hétérozygotie moyenne)
Table 3. Heterozygosity values in Postua, Cavaglià and Biella populations
Les valeurs d'hétérozygotie, d'hétérozygotie moyenne pour chaque locus et pour chaque population et la valeur de FST par locus sont indiquées dans le tableau 3. Postua présente une valeur moyenne plus basse (0.291±0.003) que celle de Cavaglià (0.329±0.002) et de Biella (0.343±0.003). La valeur de FST pour chaque locus (Tableau 3) a été calculée afin de mesurer le degré de différenciation génique entre nos populations. Cette valeur varie de 0.004 pour FXIIIB à 0,0593 pour le locus B65. La valeur moyenne de FST pour tous les locus est de 0,038.
|
Region |
Nombre d’individus |
Hétérozigositie |
FST |
|
Africa |
176 |
0.402 |
0.086 |
|
Americans |
184 |
0.381 |
0.038 |
|
Europe |
334 |
0.396 |
0.017 |
|
Sahul |
185 |
0.308 |
0.105 |
|
Southeast Asia |
359 |
0.377 |
0.067 |
|
Wester Asia |
262 |
0.414 |
0.053 |
|
Caucasus |
221 |
0.311 |
0.113 |
|
Postua-Biella-Cavaglià |
250 |
0.309 |
0.038 |
Tableau 4. Hétérozygotie moyenne et valeur de FST en Postua, Biella et Cavaglià, comparées avec d'autres régions géographiques du monde (Nasidze et al., 2001).
Table 4. Average heterozygosity and FST values for Postua Biella and Cavaglià compared with other world-wide geographic regions (Nasidze et al., 2001).
Le tableau 4 montre les valeurs d'hétérozygotie moyenne ainsi que de FST moyenne de Postua, Biella et Cavaglià, considérées globalement, avec celles se trouvant en littérature pour d'autres régions du monde (Nasidze et al., 2001).
Le valeur moyenne de FST était 0,038, laquelle met en évidence le fait que la plupart de la variance observée (96,2%) est due à des différences à l’intérieur des trois populations. Cette valeur de FST moyenne de 0,038 est plus basse que celle d'autres régions mondiales, mais elle est plus du double de la valeur connue pour l’Europe (0,017) (Nasidze et al., 2001) (Tableau 4).
Afin d'examiner les relations entre populations, nous avons construit un arbre phylogénétique Neighbor-Joining (fig. 3) obtenu en insérant non seulement les données relatives aux populations faisant l'objet de cette étude mais aussi celles obtenues par la littérature sur sept autres populations européennes (Stoneking et al., 1997).
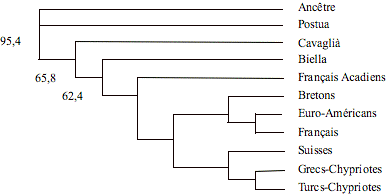
Figure 3. Arbre phylogénétique Neighbor-Joining construit avec les données de fréquence d'insertion Alu des populations étudiées, conjointement avec celles de 7 populations européennes. Les valeurs de « bootstrap » supérieures à 60% (sur un total de 1000 réplications) sont indiquées.
Figure 3. Neighbor-Joining tree of population relationships derived from Alu insertion frequencies of studied populations and other 7 European populations. Bootstrap values greater than 60% (out of 1000 replications) are indicated.
L'arbre phylogénétique a été enraciné en insérant un ancêtre hypothétique auquel a été attribuée une valeur de fréquence d'insertion égale à 0 dans chaque locus. L'on a également indiqué les valeurs de « bootstrap » supérieures à 60%, sur un total de 1000 réplications. Il est possible d'identifier deux clusters distincts, l'un représenté par les populations de Postua, Biella, Cavaglià et par les Français Acadiens, et l'autre représenté par les Français, les Bretons, les Suisses, les Euro-Américains, les Grecs et les Turcs-Chypriotes.
L'on a ensuite procédé à l’analyse de la composante principale (PC) (Cavalli-Sforza et al., 1994) fondée sur les fréquences alléliques aux six locus polymorphes étudiés (fig. 4), en comparant les populations que nous avons étudiées avec celles utilisées dans la construction de l’arbre Neighbor-Joining. Dans ce cas, les deux composantes principales représentent 86% de la variance observée (75% la première composante et 11% la seconde) et elles ont été utilisées pour déterminer les positions correspondantes des populations.
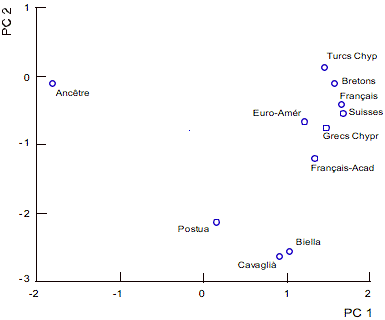
Figure 4. Graphique des deux principales composantes (PC) des fréquences alléliques à 6 locus d'insertion de séquences Alu. Les deux premières composantes principales expliquent respectivement 75% et 11% environ de la variance observée.
Figure 4. Plot of the first two principal components of the allele frequencies at the 6 Alu insertion loci. The first two principal components account for 75 and 11% respectively of the variance observed.
Dans le graphique Postua est située dans une position relativement isolée par rapport à celle des deux autres groupes, représentés respectivement par Biella-Cavaglià d'une part et par un groupe plus compact des autres populations européennes d'autre part.
Discussion
La présente étude a eu pour but de contribuer à décrire, en utilisant des marqueurs génétiques polymorphes, la structure génétique de trois populations limitrophes situées dans une région pré-alpine du Piémont oriental: Postua, Biella et Cavaglià. La population de Postua est actuellement caractérisée par des dimensions réduites (570 habitants), et elle s'est signalée, dans le passé, par un isolement partiel du point de vue géographique et socioculturel.
Les analyses biodémographiques ont mis en évidence le fait que cette population s'est distinguée par des taux élevés d'isonymie et de consanguinité dans la dernière partie du XIXe siècle et dans la première moitié du XXe siècle. Ce n'est qu'après la deuxième guerre mondiale que la courbe de l'isonymie observée ne s'écarte pas de la courbe escomptée.
L'analyse des marqueurs Alu a été effectuée sur un échantillon très représentatif de la population de Postua (102 sujets sur 570 habitants) et tous originaires de Postua depuis au moins 3 générations. Les résultats concernant les fréquences d'insertion d'éléments Alu (Tab. 2) indiquent que dans 5 locus sur 6, Postua présente les valeurs les plus basses.
Tous les locus sont polymorphes dans toutes les populations, à l’exception du locus A25 à Postua, lequel est fixé à cause de l’absence de l’élément Alu. Il s'agit là d'une donnée différente de ce qu'indique la littérature (Batzer et al. 1994) relativement à d'autres populations européennes où a été observée une gamme des valeurs de fréquence d'insertion dans le locus A25 comprise entre 0.09 et 0.160
Bien que Postua ne puisse aujourd'hui être évidemment définie comme une localité isolée, il est possible que, dans le passé, un isolement géographique relatif allié à un nombre réduit d'habitants et à un taux élevé de mariages entre consanguins, puissent avoir entraîné des fluctuations non occasionnelles des fréquences alléliques. L'un des principaux effets de l’augmentation du taux de consanguinité est la perte, à chaque génération, d'une part d'hétérozygotes au profit des homozygotes (variance de Wahlund). Comme le montre le tableau III, Postua présente une valeur d'hétérozygotie moyenne inférieure (0,291) à celle de Cavaglià et de Biella (0,329 et 0,343, respectivement). Les valeurs plus basses d'hétérozygotie moyenne observées à Postua pourraient donc être dues au nombre élevé de mariages entre consanguins ayant eu lieu au cours des périodes 1876-85 et 1926-35. Les valeurs d'hétérozygotie moyenne par locus varient de 0,056 pour A25 à 0,487 pour FXIIIB. Cette dernière valeur est proche de la valeur maximale théorique pour l’hétérozygotie de 0,5 pour un locus biallélique. Dans le locus A25 Postua ne présente pas d'individus hétérozygotes (hét = 0); Biella et Cavaglià montrent également de basses valeurs d'hétérozygotie. Pour le locus APO, les valeurs d'hétérozygotie sont également très basses dans toutes les populations étudiées. Nous avons ensuite comparé les données d'hétérozygotie moyenne de nos trois populations, considérées globalement, avec les valeurs se trouvant en littérature pour d'autres régions mondiales (Tableau 4). Les valeurs d'hétérozygotie, dans nos populations, sont plus basses que les valeurs connues pour les autres régions mondiales, y compris l'Europe, mais comparables à celles des populations du Caucase et du Sahul (Nasidze et al., 2001).
A ces données correspond cependant une valeur moyenne de FST des populations étudiées - considérées globalement - de 0,038: cette donnée peut être expliquée par le caractère strictement limitrophe des populations examinées, la valeur globale de Fst ne pouvant donc être très élevée.
Dans l’arbre Neighbor-Joining Postua occupe une position basale ; aussi Cavaglià occupe une position basale, mais cette position pourrait être la conséquence de l'exiguïté de l'échantillon analysé (53 sujets) par rapport à la population réelle (3.680 habitants). Pour confirmer cette donnée, il faudrait donc augmenter le nombre d'échantillons. La position de Biella se rapproche de celle des autres populations européennes.
La topologie de l’arbre Neighbor-Joining de la fig. 3 semble être confirmée par l’analyse de la composante principale (Fig. 4). Postua occupe une position relativement isolée, en tout cas la position la plus proche de l'ancêtre.
En conclusion, des facteurs culturels, tels que l'habitude de contracter mariage entre membres de la même communauté avec, en conséquence, une augmentation de la consanguinité entre les habitants, et un certain isolement que Postua a maintenu, dans le passé du moins, ont contribué de manière significative à déterminer la situation actuelle de cette population, peu hétérogène et caractérisée par une valeur d'hétérozygotie moyenne inférieure à celle des populations limitrophes analysées ainsi que par une plus basse fréquence d'insertions Alu dans presque tous les locus examinés.
Remerciements. Nous tenons à remercier toute la population de Postua qui a participé avec enthousiasme et disponibilité. Un grand merci, surtout, au pharmacien de Postua, M. Flavio Zanellati, qui tout mis en œuvre pour le succès de l'initiative. Merci, aussi au Professeur Gabriella Sella et au Professeur Emma Rabino Massa qui ont coordonné le projet d'étude sur la population de Postua, ainsi qu'à Mme Rosalba Guala qui a réalisé la traduction du texte en langue française.
Bibliographie
Batzer, M.A., et Deininger, P.L., 1991, A human-specific subfamily of Alu sequences. Genomics 9, 481-487;
Batzer, M.A., Stoneking, M., et Alegria-Hartman, M., 1994, African origin of human-specific polymorphic Alu insertions. Proc. Nati. Acad .Sci. USA 91, 12288-12292;
Cavalli- Sforza, L.L., Menozzi, P., et Piazza A., 1994, The History and Geography of Human genes. Princeton, NJ: Princeton University Press;
De Iasio, S., 1999, Winfamy – Speech Assistant Release 1.0. Italian C.N.R. pubblication;
Deininger, P.L., et Batzer, M.A., 1993, Evolution of retroposons. Evol. Biol. 27, 157-196;
Deininger, P.L.,et Batzer, M.A., 1999, Alu repeat and human disease. Mol. Genet. Metab. 67, 183-193;
Felsenstein ,J., 1989, PHYLIP – Phylogeny Inference Package. Cladistics, 5, 164-166;
Guo, S., et Thompson, E., 1992, Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics, 48, 361-372;
Hammer, M.A., 1994, A recent insertion of an Alu element on the Y chromosome is a useful marker for human population studies. Mol. Biol. Evol. 11, 749-761;
Kass, D.H., Aleman, C., Batzeer, M.A., et Deininger, P.L., 1994, Identification of a human specific Alu insertion in the factor XIIIB gene. Genetica 94, 1-8;
Nasidze, I., Risch, G.M., Robichaux, M, Sherry, S.T., Batzer, M.A., et Stoneking M., 2001, Alu insertion polymorphisms and the genetic structure of human populations from the Caucasus. European Journal of Human Genetics, 9, 267-272;
Okada, N., 1994, Retroposones as time marker of evolution. Proteins, Nucleic Acids, and Enzymes, 39, 2724-2735;
Ray-Engel, A. M., Carroll, M. L., Vogel, E., Garber, R. K., Nguyen, S. V., Salem, A.H., Batzer, M. A., et Deininger, P. L., 2001, Alu insertion polymorphisms far the study of human genome diversity. Genetis, 159, 279-290;
Stoneking, M., Fontius, JJ., et Clifford, SL., 1997, Alu insertion polymorphisms and human evolution: evidence for a larger population size in Africa. Genome Res., 7:1061-1071;
Ullu, E., Esposito, V., et Melli, M., 1982, Evolutionary conservation of the human 7S RNA sequences. J Mol Biol., 15, 195-201;
Walsh, P.S., Metzger, D.A., et Higuchi, R., 1991, Chelex 100 as a Medium for simple extraction of DNA for PCR-based typing from forensic material. BioTechniques, 10, 506-513;
Weiner, A.M., Deininger, P.L., et Efstratiadis, A., 1986, Nonviral retroposons: genes, pseudogenes, and transposable elements generated by the reverse flow for genetic information. Annu. Rev. Biochem., 55, 631-661.