
Colloques du Groupement des Anthropologistes de Langue Française (GALF)
Díaz N., Solórzano E., Montiel, R., García, C., Yañez C., Malgosa, A., 2004, Determination genétique de l’individu Néolithique de Segudet (Ordino), les restes humains les plus anciens d’Andorre. Antropo, 7, 39-44. www.didac.ehu.es/antropo
Determination genétique de l’individu Néolithique de Segudet (Ordino), les restes humains les plus anciens d’Andorre
Genetic determination of the Neolithic individual of Segudet (Ordino), these are the most ancient human remains found in Andorra
Díaz N.1,4, Solórzano E.1,4, Montiel, R.1,2, García, C.1,4, Yañez C.3, Malgosa, A.1
1Unitat d'Antropologia. Dept. Biologia Animal, Vegetal i Ecologia, Universitat Autònoma de Barcelona. España. E-mail: Assumpcio.Malgosa@uab.es
2 Departamento de Biologia - CIRN, Universidade dos Açores. Portugal.
3 Àrea de Recerca Històrica del Patrimoni Cultural d’Andorra.
4 Facultad de Odontología. Universidad de Los Andes. Venezuela.
Correspondance: Assumpció Malgosa. Unitat d'Antropologia. Dept. Biologia Animal, Vegetal i Ecologia, Universitat Autònoma de Barcelona. España. E-mail: Assumpcio.Malgosa@uab.es
Mots-clés: ADN mitochondrial, RFLP’s, Néolithique, Andorre.
Keywords: DNA mitochondrial, RFLP’s , Neolithic, Andorra.
Resumé
Les restes humains analysés proviennent d’une tombe préhistorique découverte à Segudet dans la paroisse d’Ordino. Le squelette appartient à un individu adulte, de sexe féminin, dont l’age au décès peut être évalué à 30/35 ans. Le résultat de l'analyse par la méthode de C14 (BETA - 160374) a permis de déterminer que ces restes humains dataient de 5350+/- 40 BP. Par conséquent, ils sont les plus anciens mis à jour en Andorre et cet individu appartient à la période du Néolithique ancien (période épicardial).
La détermination génétique de l’ADN mitochondrial par RFLP’s et la séquence d’un fragment de la région de contrôle ont permis de classer l’individu dans l’haplogroupe européenne K. Une interprétation portant sur le groupe humain est irréalisable, mais la relation entre l’ancienneté de cet haplogroupe et la situation stratégique d’Andorre comme voie d’accès à la Péninsule Ibérique est remarquable.
Abstract
Human remains analyzed in this study come from a prehistoric burial from Segudet, Ordino, Andorra. The skeleton was diagnosed as a female of between 30-35 years of age at death. C14 analysis results in 5350±40 BP; this date and the funerary furnishings found in the burial set it in the ancient Neolithic Age, specifically in the Epicardial period. These are the most ancient human remains found in Andorra.
Genetic determination was performed by mitochondrial haplogroup analysis and sequencing the control region. Both analysis enable us to classify this individual within the European haplogroup K. Although population interpretation cannot be carried out, it is interesting to notice the antiquity of this haplogroup in Europe and the strategic location of the country of Andorra as a pass to enter the Iberian Peninsula.
Introduction
Les restes humains analysés proviennent d’une tombe préhistorique découverte à Segudet dans la paroisse d’Ordino à une altitude de 1324 m. Cet individu appartient á la période du néolithique ancien (période épicardial, 4300 et 4050 av-JC). Ces restes, déposés pour étude au laboratoire de Paleoanthropologie de la Unitat d'Antropologia, Dept. Biologia Animal, Vegetal i Ecologia, Universitat Autònoma de Barcelona, ont été identifiés sous le sigle S-2001.
Le squelette appartient à un individu adulte, de sexe féminin, dont l’âge au décès est évalué entre 30 et 35 ans. D’autre part, le résultat de l'analyse par la méthode de C14 (BETA - 160374) a permis de déterminer que ces restes humains, datés de 5350+/- 40 BP, étaient les plus anciens mis à jour en Andorre (Yañez et al., 2003) (fig. 1).
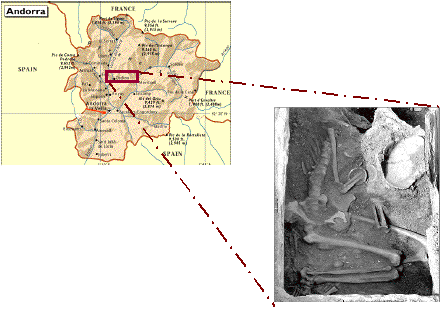
Figure 1. Localisation géographique du gisement néolithique de Segudet à Ordino et détail de la tombe.
On peut noter la richesse du trousseau (p.e. bracelets).
Figure 1. Geographic location of the Neolithic site of Segudet, Ordino, and photograph of the burial.
Wealth of the furnishings can be noted (ex. bracelets).
Matériel et méthode
Pour obtenir de l'ADN à partir des restes anciens il a fallu modifié les techniques employées par la biologie moléculaire de sorte à les adapter aux conditions de conservation de ce type d'échantillon pour lequel l’ADN est fréquemment trop dégradé pour permettre d’appliquer des procédés trop délicats (Krings et al., 1997). Il faut également tenir compte que le spécimen est susceptible d’avoir été contaminé par l’ADN moderne (Montiel et al., 2000, 2001).
L'analyse moléculaire a été effectuée suivant la méthode employée par notre laboratoire. Celui-ci consiste, en l’obtention d’ADN à partir de pièces dentaires et suit les étapes suivantes : extraction organique, concentration et purification de l’ADN obtenu (Montiel, 2001). Pour effectuer ce travail, il est nécessaire d’employer des échantillons en excellent état (Fig 2), c’est pourquoi notre choix c’est porté sur la pièce dentaire 33 (canine supérieure droit).
Figure 2. L’ADN a été obtenue de la chambre pulpaire des dents de l’individu S-2001.
Figure 2. ADN was obtained from the pulp chamber of teeth of the S-2001 individual.
Pour la caractérisation génétique de l’individu nous avons employé l’ADN mitochondrial, en combinant les résultats de deux systèmes: les polymorphismes de la longueur des fragments de restriction (RFLP's) décrits pour les populations européennes et l’analyse des séquences d’un fragment de 230 pb de la région de control (HVI).
Resultats et discussion
L’analyse des 10 points polymorphiques de restriction (Torroni et al., 1996) avec l’utilisation des primers qui amplifient de petits fragments (Garcia, 2002a): la séquence de 180 pb, obtenue a partir de l’amplification d’un fragment de la région HVI, nous a permis de déterminer que l’individu étudié appartient au haplogroupe européen K (fig. 3 table 1 et 2).
Six des haplogroupes Européens (H, I, J, K, T et W) sont fortement confinés a des populations Européennes (Torroni et al., 1994, 1996) et leur arrivée est postérieure à celle des premiers caucasoïdes qui sont déjà séparés génétiquement des ancêtres Africains modernes et Asiatiques (Ozawa 1995, Torroni et al., 1996; Passarino et al., 1998; Macaulay et al., 1999). Ce haplogroupe, caractérisé par un âge prévu entre 12.900 et 18.300 ans, semble être l’un des plus anciens (Richards et al., 2000). D’autre part, sur un échantillonnage des populations européennes contemporaines, l’haplogroupe K présente une proportion variant de 4.4 à 16.6% (Table 3), la population autochtone d’Andorre présentant la fréquence la plus élevée.
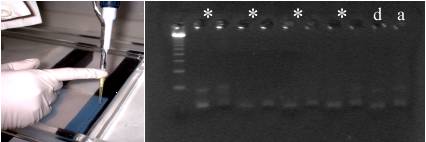
Figure 3. Electrophoresis of restriction with HaeII enzyme (-9052).
*other samples, d = digestion, a = amplification.
|
|
I-X |
V |
H |
W |
K |
Eur |
U |
J |
T |
|
|
-1715 DdeI |
- 4577 NlaIII |
-7025 AluI |
- 8994 Hae III |
- 9052 HaeII |
+/-10394 DdeI |
+ 12308 HinfI |
- 13704 Bst0I |
+ 15606 AluI |
|
Segudet |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
- |
|
|
No I-X |
No V |
No H |
No W |
K |
No Eur |
U |
No J |
No T |
Table 1. Définition des haplogroupes sur la base de la présence (+) ou l’absence (-) des lieux de restriction (Torroni et al., 1996).
Table 1. Definition of haplogroups based on presence (+) or absence (-) of restriction sites (Torroni et al., 1996).
|
|
1 6 2 2 0 |
1 6 2 2 4 |
1 6 2 2 9 |
1 6 2 4 1 |
1 6 2 4 4 |
1 6 2 5 7 |
1 6 2 6 0 |
1 6 2 6 1 |
1 6 2 7 4 |
1 6 2 8 5 |
1 6 2 9 0 |
1 6 3 1 1 |
1 6 3 1 6 |
1 6 3 2 5 |
1 6 3 4 4 |
1 6 3 5 1 |
1 6 3 5 7 |
1 6 3 6 2 |
1 6 3 8 9 |
1 6 4 0 0 |
HVI |
RFLP |
|
Anderson |
A |
T |
T |
A |
G |
C |
C |
C |
G |
A |
C |
T |
A |
T |
C |
A |
T |
T |
G |
C |
|
|
|
Segudet |
. |
C |
. |
. |
. |
. |
. |
. |
. |
. |
. |
C |
. |
. |
. |
. |
. |
. |
. |
. |
Haplo K |
Haplo K |
Table 2. Séquence de la région HVI de l’individu S-2001. Le point indique la présence du même nucléotide qu’il y a dans la séquence de référence (CRS).
Table 2. HVI sequence of the individual S-2001. Points indicate the same nucleotide as in the reference sequence (CRS). Mutations have been pointed out with blue colour.
|
Haplogroupes |
Pays Basque N=156 |
Catalogne* N=90 |
Andorre** N=99 |
|
X |
1.2 |
2.2 |
0.0 |
|
V |
10.8 |
3.3 |
3.5 |
|
K |
4.4 |
5.6 |
16.6 |
|
H |
58.9 |
44.4 |
51.8 |
|
U |
14.1 |
11.1 |
22.4 |
|
T |
5.1 |
13.3 |
9.4 |
|
W |
- |
0.0 |
0.0 |
|
J |
2.5 |
13.3 |
4.7 |
|
I |
- |
0.0 |
7.1 |
|
Autre |
2.5 |
- |
- |
Table 3. Fréquences des haplogroupes dans des populations ibériques actuelles.
Source: Bertranpetit et al., 1995 et Corte-Real et al., 1996 ; * Montiel, 2001 ; * Anglés et al., 2003.
Table 3. Haplogroup frequencies in present Iberian populations.
Source: Bertranpetit et al 1995 et Corte-Real et al., 1996 ; * Montiel, 2001 ; ** Anglés et al., 2002.
La signification de ces résultats doit être évaluée dans le contexte géographique, temporel et culturel de la découverte archéologique. De plus, il est nécessaire de prendre en compte que le lieu de la découverte est situé sur un axe de passage traversant les Pyrénées qui présente des caractères comparables à ceux enregistrés pour d’autres gisements néolithiques européens.
Dans le contexte ancien de la Péninsule Ibérique, cet haplogroupe est présent dans les populations néolithiques du Pays Basque (Izaguirre, 1999). En Catalogne, il n’apparaît ni dans les séries néolithique, ni dans celles du Bronze du Mas d’en Boixos (Garcia et al., 2002), alors qu’il est présent dans celle de la population médiévale de la Plaça Vella (Table 4). C’est pour cette raison que sa présence dans le groupe des populations anciennes ayant pu entrer dans la Péninsule n’est pas surprenante et qu’elle n’est pas non plus exceptionnelle dans la population néolithique d’Andorre.
|
Haplogroupes |
SJAPL* N=61 |
Longar* N=27 |
Pico Ramos* N=24 |
Plaça Vella** N=24 |
Mas d´Boixos*** (Néolithique) N=5 |
Mas d´Boixos*** (Bronze) N=12 |
|
X |
0.0 |
0.0 |
0.0 |
- |
- |
- |
|
I,K,J |
39.4 |
22.2 |
33.3 |
33.4 |
40.0 |
8.3 |
|
V |
0.0 |
0.0 |
0.0 |
12.5 |
- |
- |
|
H |
37.7 |
40.7 |
37.5 |
33.3 |
20.0 |
75.0 |
|
U |
18.0 |
14.8 |
12.5 |
8.3 |
- |
- |
|
T |
4.9 |
14.9 |
16.6 |
8.3 |
- |
- |
|
W |
0.0 |
0.0 |
0.0 |
- |
- |
- |
|
? |
- |
- |
- |
4.2 |
20.0 |
16.6 |
|
H,X,V,U,W? |
- |
- |
- |
- |
20.0 |
- |
Table 4. Fréquences des haplogroupes dans des séries anciennes.
Source : * Izaguirre, 1998 ** Montiel, 2001 ; *** Garcia, 2002.
Table 4. Haplogroup frequencies in ancient Iberian populations.
Source: * Izaguirre, 1998, ** Montiel, 2001; *** Garcia, 2002.
Conclusions
Ces restes anciens sont les premiers restes analysés du point de vue génétique en Andorre.
La combinaison des résultats des deux systèmes: RFLP's et l’analyse de la séquence de la région HVI, nous indiquent que l’individu S-2001, appartient au haplogroupe européen K.
On ne peut pas affirmer la relation génétique avec les autres gisements néolithiques catalans, dont les données sont insuffisantes alors que la relation culturelle (le trousseau) est indiscutable.
Comme ce haplogroupe a également été mis en lumière pour les populations du même âge du Pays Basque, un lien peut perçu en se fondant sur la concomitance de leur situation stratégique dans les Pyrénées.
Bien peu d’analyses paleogénétiques ont été effectuées dans notre contexte géographique, il est par conséquent évident que de nombreuses études seront à réaliser afin de définir les relations entre groupes de population.
Reconnaissance: Àrea de Recerca Històrica del Patrimoni Cultural d’Andorra.
Bibliografie
Anderson, S., A.T. Bankier, B.G. Barrell, M.H.L. Bruijn, A.R. Coulson, J. Drovin, I.C. Eperon, D.P. Nierlich, B.A. Roe, F. Sanger, P.H. Schreier, A.J.H. Smith, R. Staden y I.G. Young. 1981. Sequence and organization of the human mitochondrial genome. Nature. 290: 457-464.
Anglés, N., Montiel, R., Nogués, R.M., Backes, Ch., Larrouy, G., Cherkaoui, M., Malgosa, A., Aluja, M.P., 2003, Haplogrupos mitocondriales en tres poblaciones caucasoides. Dans: Antropología y Biodiversidad, edité par M.P. Aluja, A. Malgosa, R.M. Nogués, (Barcelona: Edcs. Bellaterra) vol 2, pp. 26-35
Bertranpetit, J., Sala, J., Calafell, F., Underhill, P.A., Moral, P., Comas, D., 1995, Human mitochondrial DNA variation and the origin of Basques. Ann. Hum. Genet., 59, 63-81
Côrte-Real, H.; Macaulay, V.; Richards, M.; Hariti, G.; Issad, A.; Cambon-Thomsen, A.; Papiha, S.; Bertranpetit,J y Sykes, B.1996. Genetic diversity in the Iberian Peninsula determined from mitochondrial sequence analysis. Ann. Hum. Genet. 60: 331-350.
García, C, 2002, Análisis de los Haplogrupos del DNAmt en restos dentales del Neolítico Catalán. MS Thesis, Universitat Autònoma de Barcelona.
García, C., Montiel, R., Díaz, N., Malgosa, A., 2002, Mitochondrial aDNA variability of a Catalonian site from Neolithic and Bronze Age. Congress of the European Anthropological Association. Zagreb 29/09-3/10/2002
Izaguirre, N., 1998, Aplicación de la biología molecular en el estudio antropológico de las poblaciones humanas del pasado: análisis del ADN mitocondrial. Ph.D. Thesis. Euskal Herriko Unibersitatea.
Krings, M., Stone, A., Schimtz, R.W., Krainitzki, H., Stoneking, M., Paabo, S., 1997, Neanderthal DNA sequences and the origin of modern humans. Cell, 90, 19-30
Macaulay, V., Richards, M., Sikes, B., 1999, Mitochondrial DNA recombination – no need to panic. Proc. R. Soc. Lond. B., 266, 2037-2039.
Montiel, R., 2001, Estudio diacrónico de la Variabilidad del DNA mitocondrial en Población catalana. Ph,D. Thesis, Universitat Autònoma de Barcelona.
Montiel, R., Malgosa, A., Francalacci, P., 2000, Autentificación del ADN antiguo. Dans: Investigaciones en Biodiversidad, edité par: T.A. Varela (Santiago de Compostela: Ed.Universidad de Santiago de Compostela) pp. 762- 772
Montiel, R., Malgosa, A., Francalacci, P., 2001, Authenticating ancient human mitochondrial DNA. Human Biology; 73(5), 689-713.
Ozawa T., 1995, Mechanism of somatic mitochondrial DNA mutations associated with age and diseases. Biochim Biophys Acta 1271: 177-189.
Passarino, G., Semino, O., Quintana-Murci, L., Excoffier, L., Hammer, M., Santachiara-Benerecetti, A.S., 1998, Different genetic components in the Ethipian population, identified by mtDNA and Y-chromosme polymorphism. Am. J. Hum. Genet 62(2), 420-434.
Richards, M., Macaulay, V., Hickey, E., Vega, E., Sykes, B., Guida, V., Rengo, C., Sellito, D., Cruciani, F., Kivisild, T., Villems, R., Thomas M., Rychkov, S., Rychkov, O., Rychokv, Y., Gölge, M., Dimitrov, D., Hill, E., Bradley, D., Romano, V., Calì, F., Vona G., Demaine, A., Papiha, S., Triantaphyllidis, C., Stefanescu, G., Hatina, J., Belledi, M., Di Rienzo, A., Novelletto, Andrea., Oppenheim, A., NØrby. S., Al-Zaheri, N., Santachiara-Benereccetti, S., Scozzari, R., Torroni, A., Bandelt, HJ. 2000, Tracing European Founder Lineages in the Near Eastern mtDNA Pool. Am. J. Hum. Genet., 67, 1251-1276.
Torroni, A., Lott, M.T., Cabell, M.F., Chen, Y.S., Lavergne, L., Wallace, D.C., 1994, mtDNA and the origin of Caucasians: identification of ancient Caucasian-specific haplogroups, one of which is prone to a recurrent somatic duplication in the D-loop region. Am. J. Hum. Genet., 55, 760-776.
Torroni, A., Huoponen, K., Francalacci, P., Petrozzi, M., Morelli, L., Scozzari, R., Obinu, D., Savontaus, M.L., Wallace, D.C., 1996, Classification of European mtDNAs from an analysis of three European populations. Genetics 144, 1835-1850.
Yáñez, C., Malgosa, A., Burjachs, F., Díaz,
N., García, C., Isidro, A., Juan, J., Matamala, J., 2002, El món funerari al
final del V mil.leni a Andorra: La tomba de Segudet (Ordino). Cypsela,
* *
* *