
Colloques du Groupement des
Anthropologistes de Langue Française, GALF)
Sabir B., Cherkaoui M., Baali A., Hachri H., Lemaire O., Dugoujon J.M.,
2004, Les
dermatoglyphes digitaux et les groupes sanguins ABO, Rhésus et Kell dans une
population Berbère du Haut Atlas de Marrakech.
Antropo, 7, 211-221. www.didac.ehu.es/antropo
Les dermatoglyphes digitaux et les groupes
sanguins ABO, Rhésus et Kell dans une population Berbère du Haut Atlas de
Marrakech
The digital dermatoglyphics and ABO, Rhesus and Kell blood groups in a Berber population of the High Atlas of Marrakech
Sabir B.1,
Cherkaoui M.2, Baali A.2, Hachri H.2, Lemaire O.3, Dugoujon J.M.3
1 Hôpital d’Asni, Délégation du
Haouz, Ministère de la Santé, Maroc. E-mail: b.sabir@ucam.ac.ma
2 Laboratoire d’Ecologie Humaine,
Faculté des Sciences Semlalia, Université Cadi Ayyad, Marrakech, Maroc
3 UMR 8555 CNRS, Centre d’Anthropologie, Toulouse, France
Mots clés: Dermatoglyphes, systèmes sanguins, Berbères.
Key words: Dermatoglyphics, blood groups, Berber.
Résumé
Dans cette étude, nous présentons les résultats de l’analyse des
dermatoglyphes digitaux et des systèmes de groupe sanguin (ABO, Rhésus et Kell)
dans une population saine de 120 personnes d’origine berbère (tribu des
Rhiraya, dans le Haut Atlas Occidental). L’enquête a été réalisée en 2002
à Asni (Province du Haouz, Wilaya de Marrakech). Nous avons observé 50.2%
de boucles ulnaires, 40.6% de tourbillons, 7.7% d'arcs et 1.5% de boucles
radiales. Les fréquences des groupes sanguins ABO sont de 40.7 % pour le groupe
O, 37.2 % pour le groupe A, 19.5% pour le groupe B et 2.7 % pour le groupe AB.
Pour le système Rhésus, 89.3% des sujets sont Rh + et 10.7% Rh - ; quant
au système Kell, 5.4% sont K + et 94.6% K -. Concernant les figures digitales, l’analyse montre que notre
population se distingue des autres populations marocaines et méditerranéennes.
Elle se rattache plus au groupe des populations du sud de la méditerranée
(arabes et berbères) qu’au groupe des populations du nord (Espagne, France et
Italie). L’analyse en composantes principales des
fréquences des allèles de groupe sanguin confirme cette position intermédiaire
entre les populations arabes et du Moyen Orient et les populations
Sud-européennes.
Abstract
In this survey, we present results of
the analysis of the digital dermatoglyphics and the blood systems (ABO,
Rhesus and Kell) of a sample of
120 individuals of origin Berber Rhiraya (High Atlas Western, Marrakech),
gotten at the time of a country of sampling achieved in 2002 to the hospital of
Asni (Province of the Haouz, Wilaya of Marrakech). .The frequencies of the digital figures obtained in
this population are respectively 50.2% of the ulnar loops, 40.6% of the whorls,
7.7% of the archs and 1.5% of the radial loops. The
frequencies of ABO blood types recorded are 40.7% for the O group , 37.2% for
the A group, 19.5% for the B group and 2.7% for the AB group. The
Rhesus System is represented by 89.3% of Rh+ and 10.7% of Rh- and the system Kell by 5.4% K+ and 94.6%
K-. The analysis of affinities of
the digital figures between the population object of the study and those of the
Mediterranean by the Euclidean distances show that this one presents
characteristics which differentiates it from the other populations. One can
note however, that conversely with other Moroccan populations analysed, the one
becomes attached more to the group of the populations of the south of the
Mediterranean (Arab and Berber) than to the group of the populations of north (Spain,
France and Italy) .The analysis in Main Component of
blood type frequencies gotten confirms this intermediate position of our
population between the Arabian and middle oriental populations and the European
populations of the South.
Introduction
Depuis la protohistoire, le pourtour méditerranéen connaît un mouvement
ininterrompu d’hommes et d’idées
brassant ses peuples et ses cultures. Tous les peuples de la Méditerranée
(Phéniciens, Romains, Vandales, Byzantins, Arabes, Turcs et Européens) ont traversé
l'Afrique du Nord et ont contribué à enrichir culturellement cette vaste
région. Dans le passé, l’Afrique du Nord a toujours été occupé par des
Berbères. Les historiens grecs et latins les nommaient sous des noms divers
(Garamantes, Maures, Numides, Gétules, Nasamons, Psyles…) ; les nouveaux
arrivants, toujours minoritaires, étaient assimilés dans ce fond berbère
autochtone (Camps, 1980).
Le terme Berbère désigne l’ensemble de populations qui ont parlé ou
parlent encore des dialectes apparentés à un fond commun « la langue
berbère ». Les Berbères ont occupé de vastes territoires en Afrique du
Nord pendant des milliers d’années. Ils se sont surtout maintenus dans les
zones montagneuses (Rif et Atlas marocains, Aurès et Kabylie en Algérie), dans
des régions isolées (Matmata et Djerba en Tunisie, oasis de Siwa en Egypte), ou
dans le cas des Touaregs nomades en zone désertique (Mali, Mauritanie, Tchad,
Sénégal et Burkina-Faso).
La question de
l’origine des Berbères s’est posée tout le long de l’histoire de l’Afrique du
Nord. Elle a suscité d’énormes débats et d’innombrables théories. Certains
auteurs croient qu’ils sont venus d’Europe. Mais la plupart pense qu’ils sont
les descendants d’une population autochtone apparue in situ en Afrique du Nord,
de culture paléolithique Ibéro-maurusienne (-16 000 ans), puis mésolithique
capsienne. Enfin, d’autres les considèrent issus de populations orientales
ayant migré dans cette région durant la transition néolithique (- 9 500, - 7
000 ans).
Notre étude,
réalisée dans le cadre du projet CNRS et de l’ESF (EUROCORES): «Le berbère
et les Berbères: diversités linguistique et génétique», a pour objectif
d’enrichir la base de données anthropogénétiques sur les Berbères. Nous
présentons ici les résultats des
recherches (dermatoglyphes et systèmes de groupes sanguins) effectuées sur une population
berbère de la région d’Asni. La diversité génétique sera comparée à celle
d’autres populations berbères et arabes d’Afrique du Nord, ainsi que de
l’espace méditerranéen.
Matériel et méthodes
L’échantillonnage
a été réalisé sur une population issue de différents villages localisés dans le Haut Atlas
occidental, dans la commune d'Asni (Province du Haouz, Wilaya Marrakech),
située à environ 50 Km de la ville de Marrakech. Au niveau ethnique, la
population est très homogène; elle appartient à la
tribu Rhiraya, du grand groupe Masmouda, établi
dans le Haut Atlas avant le XIIème siècle
(Pascon, 1977). Les prélèvements sanguins ont
été effectués sur des volontaires (au consentement éclairé), lors d’une
campagne réalisée par l’équipe participante au projet avec l’aide du personnel
médical de l’hôpital d’Asni. Les individus répondaient à 3 critères : ils
parlent le Berbère, ils sont originaires de la région et ils sont non
apparentés. Pour les dermatoglyphes, nous
avons utilisé le procédé qui consiste à enduire la surface ridée d’encre.
L’extrémité distale de chacun des dix doigts est roulée sur le papier de façon
à obtenir la totalité du dessin. Notre analyse a porté sur 100 sujets
adultes (50 hommes et 50 femmes). Pour les échantillons sanguins, chaque donneur a été prélevé par ponction veineuse (sur des tubes
héparinés et EDTA). Les prélèvements ont été transférés au laboratoire d’Ecologie Humaine où
ils ont été conservés à 4°C, avant leur acheminement vers l’UMR 8555 du CNRS
pour analyses. Cent treize échantillons ont été testés. L’analyse des
dermatoglyphes a été réalisée selon la méthodologie de Cummins et Midlo (1961)
et de Penrose (1968). Les tests statistiques classiques: ![]() et Kolmogrov-Smirnov (K. S.) ont été utilisés
pour mesurer les différences mono-factorielles. Un dendrogramme (dérivé de la
matrice des
et Kolmogrov-Smirnov (K. S.) ont été utilisés
pour mesurer les différences mono-factorielles. Un dendrogramme (dérivé de la
matrice des
distances euclidiennes) pour les fréquences des figures digitales et une analyse en composante principale obtenue à partir de la matrice des fréquences des groupes sanguins ont été réalisés. Le traitement séparé des données des dermatoglyphes digitaux et des groupes sanguins est dû à leur déterminisme génétique différent : pour les groupes sanguins, le déterminisme génétique est mendélien et on exprime les différences entre populations en terme de fréquences alléliques et génotypiques, alors que pour les dermatoglyphes le déterminisme est polygénique (leur forte héritabilité et leur stabilité font d’eux de bons marqueurs génétiques) et les différences sont exprimées en terme de phénotypes. La base de données bibliographiques ayant servie pour l’étude comparative figure dans les tableaux 1 et 2. Les analyses statistiques ont été réalisées à l’aide du logiciel SPSS 10.0 pour Windows.
|
Population |
Code |
Pays |
Références |
|
Kabyles |
Algé1 |
Algérie |
Chamla, 1961 |
|
Algérie (total) |
Algé2 |
Algérie |
Chamla, 1961 |
|
Chypre |
Chyp1 |
Chypre |
Plato, 1970 |
|
Iles Baléares (Minorque) |
Espa1 |
Espagne |
Moreno et Pons, 1985 |
|
Andalousie |
Espa2 |
Espagne |
Oyhenart, 1985; Luna et Pons, 1987 |
|
Alcarria |
Espa3 |
Espagne |
Portabales, 1983 |
|
Sierra del Gredos |
Espa4 |
Espagne |
Fuster et Cabello, 1985 |
|
Catalogne (Barcelone) |
Espa5 |
Espagne |
Pons, 1952 |
|
Pays Basque |
Espa6 |
Espagne |
Arrieta, 1985 |
|
Asturies |
Espa7 |
Espagne |
Egocheaga, 1973 |
|
Galice |
Espa8 |
Espagne |
Oyhenart, 1983 |
|
France |
Fran |
France |
Gessain et Gessain, 1956 |
|
Grèce |
Grèc |
Grèce |
Weninger et al., 1974; Roberts et al., 1965 |
|
Bologne |
Ital1 |
Italie |
Gualdi-Russo et al., 1982 |
|
Sardaigne |
Ital2 |
Italie |
Floris et Sanna, 1986; Vrydagh-Laoureux, 1966 |
|
Vallouise |
Ital3 |
Italie |
Salis et al., 2001 |
|
Liban |
Liba |
Liban |
Naffah, 1974 |
|
Berbères (Libye) |
Liby |
Libye |
Falco, 1917 |
|
Arabes (Libye) |
Liby |
Libye |
Falco, 1917 |
|
Malte |
Malt |
Malte |
Bozicevic et al., 1993 |
|
Berbères de la région d’Asni |
Maro1 |
Maroc |
présente étude |
|
Berbères du Moyen Atlas |
Maro2 |
Maroc |
Harich et al., 2002 |
|
Marocains de Belgique |
Maro3 |
Maroc |
Vrydagh-Laoureux, 1979 |
|
Arabes méridionaux (Doukalla) |
Maro4 |
Maroc |
Kandil et al., 1998 |
|
Beni Methar |
Maro5 |
Maroc |
Gessain, 1957 |
|
Portugal |
Port |
Portugal |
Cunha et Abreu, 1954 |
|
Bédouins Rwala (Syrie) |
Syri |
Syrie |
Shanklin et al., 1937 |
|
Tunisie (Tunis) |
Tun |
Tunisie |
Chamla, 1973 |
Tableau 1. Base de données pour l’analyse des dermatoglyphes.
|
Population |
Code |
Pays |
Source de données |
|
Berbères (Tizi-Ouzou) |
Alg1 |
Algérie |
Ruffié et al., 1962; (1) |
|
Algérie (Alger) |
Alg2 |
Algérie |
Ruffié et al., 1962 ; (1) |
|
Algerie - Zemmora (les Flittas) |
Alg3 |
Algérie |
Auzas, 1957 |
|
Berbères (Tlemcen) |
Alg4 |
Algérie |
Ruffié et al., 1962 ; (1) |
|
Oran |
Alg5 |
Algérie |
Auzas, 1957 |
|
Touareg de l'Aïr |
alg6 |
Algérie |
Barnicot et al., 1954 ; (1) |
|
Arabie Saoudite (tribus de l’est) |
ara1 |
Arabie Saoudite |
Maranjiani et al., 1966 ; (3) |
|
Arabie Saoudite (tribus de l’ouest) |
ara2 |
Arabie Saoudite |
Saha et al., 1980 |
|
Chypre |
chy1 |
Chypre |
Poumpouridou et al., 1995 |
|
Egypte (Le Caire) |
egy1 |
Egypte |
Matta, 1937 ; (1) |
|
Egypte (Sinai) |
egy2 |
Egypte |
Bonne et al., 1971 ; (1) |
|
Espagne (Minorque) |
esp1 |
Espagne |
Moral, 1986 |
|
Espagne (Galice) |
esp2 |
Espagne |
Gauch et al., 1952 ; (2) |
|
Espagne (Alpajurras) |
esp3 |
Espagne |
Fernandez et al., 1999 |
|
Centre de l'Espagne |
esp4 |
Espagne |
Colino, 1978 |
|
Espagne (Catalogne) |
esp5 |
Espagne |
Moreno et Moral, 1983 |
|
Espagne (Basques) |
esp6 |
Espagne |
Manzano et al., 1996 |
|
France (Corse) |
fra1 |
France |
Memmi, 1999 |
|
Sud de la France |
fra2 |
France |
Kherummian, 1961 ; (1) |
|
Grèce continentale |
gre1 |
Grèce |
Tsiakalos et al 1980 |
|
Grèce (Plait) |
gre2 |
Grèce |
Tills et al 1983 ; (3) |
|
Grèce (Crète) |
gre3 |
Grèce |
Branicot et al., 1965 ; (1) |
|
Italie (Sardaigne) |
ita1 |
Italie |
Moral et al ., 1994 |
|
Italie du Sud |
ita2 |
Italie |
Piazza et al., 1989 |
|
Italie (Sicile) |
ita3 |
Italie |
Vona et al., 1998 |
|
Italie (Lazio) |
ita4 |
Italie |
Piazza et al., 1989 |
|
Italie du Nord |
ita5 |
Italie |
Piazza et al., 1989 |
|
Jordanie (Bédouins) |
jor1 |
Jordanie |
Saha et al., 1986 ; (3) |
|
Jordanie |
jor2 |
Jordanie |
Nabulsi et al., 1997 |
|
Liban |
lib1 |
Liban |
Ruffié et Taleb, 1965 ; (1) |
|
Libye |
lyb1 |
Libye |
Walter et al., 1975 ; (3) |
Malte
|
mal1 |
Malte |
Ikin, 1963 ; (1) |
Berbères (Asni)
|
mar1 |
Maroc |
présente étude |
|
Berbères (Ouarzazate) |
mar2 |
Maroc |
Errahaoui, 2002 |
|
Berberes (Ait Haddidu) |
mar3 |
Maroc |
Johnson et al., 1963 |
|
Berbères du Moyen Atlas |
mar4 |
Maroc |
Harich, 2002 |
|
Haratin: Ait Yazza (Ait Atta) |
mar5 |
Maroc |
Mechali et al., 1957 |
|
Haratin: Ait Khalifa (Ait Atta) |
mar6 |
Maroc |
Mechali et al.,1957 |
|
Haratin (Ksar Glagla) |
mar7 |
Maroc |
Mechali et al., 1957 |
|
Haratin: Ait Ayache (Ait Izdek) |
mar8 |
Maroc |
Mechali et al., 1957 |
|
Arabes méridionaux (Doukala) |
mar9 |
Maroc |
Kandil, 1999 |
|
Maroc: Moulay Driss |
mar10 |
Maroc |
Méchali, 1955 in Harich, 2002 |
|
Terra de bouro |
por1 |
Portugal |
Cruz et al., 1973 ; (3) |
|
Tunisie (global) |
tun1 |
Tunisie |
Gherbi et al., 1965 ; (1) |
|
Turquie |
tur1 |
Turquie |
Mourant, 1983 in Solignac et al., 1995 |
|
Yémen |
yem1 |
Yémen |
Tills et al., 1983 ; (3) |
|
Yémen - Habbanite (Juifs) |
yem2 |
Yémen |
Bonné, 1970
|
Tableau
2. Base de données pour l’analyse
des Groupes sanguins. (1): cité par Mourant et al., 1976 ;
(2) : cité par Tills et al., 1983 ; (3) : cité par Roychoudhury
et Nei, 1988 ; (4) : cité par Moral, 1986
Résultats
I/
Distributions phénotypiques et alléliques
I-1/ Fréquence des figures digitales dans
la population d’Asni
Les fréquences des figures digitales
sont de 50.2% pour les boucles ulnaires, de 40.6% pour les tourbillons,
7.7% pour les arcs et de 1.5% pour les boucles radiales. On note plus de
boucles radiales et moins d’arcs chez les hommes que chez les femmes (tableau
3). Chez les hommes, les arcs plats sont surtout fréquents au niveau de DIII et
GIII, les arcs en tente au niveau du GII, les boucles ulnaires aux doigts V,
les doubles boucles aux doigts I des deux mains et les tourbillons au GIV. Les
boucles radiales sont rares chez les hommes. Chez les femmes, les arcs ont une
fréquence maximale aux DII et GII, les boucles radiales aux GII et GIII, les
boucles ulnaires aux DIII et GV, les doubles boucles aux doigts I et les
tourbillons aux doigts IV. Il n’y a pas de différence statistiquement
significative entre les deux sexes. Pour la symétrie bilatérale, on note une
différence statistiquement significative entre la main droite et la main
gauche. Cette différence est due essentiellement aux DIII et GIII. Pour les
autres doigts, la symétrie est parfaitement respectée.
|
Hommes |
|||||||||||||||
|
|
DI |
DII |
DIII |
DIV |
DV |
D |
GI |
GII |
GIII |
GIV |
GV |
G |
D+G |
||
|
Total arcs |
2 |
10 |
10 |
4 |
6 |
6 |
4 |
2 |
8 |
2 |
6 |
6.4 |
6.2 |
||
|
Arcs plats |
2 |
6 |
8 |
4 |
6 |
4.8 |
4 |
6 |
8 |
2 |
4 |
4.8 |
4.8 |
||
|
Arcs en tente |
0 |
4 |
2 |
0 |
0 |
1.2 |
0 |
6 |
0 |
0 |
2 |
1.6 |
1.4 |
||
|
Boucles ulnaires |
30 |
36 |
54 |
46 |
76 |
48.4 |
44 |
38 |
50 |
46 |
82 |
52 |
50.2 |
||
|
Boucles radiales |
0 |
4 |
0 |
0 |
0 |
0.8 |
0 |
14 |
0 |
2 |
0 |
3.2 |
2 |
||
|
Total tourbillons |
68 |
50 |
36 |
50 |
18 |
44.8 |
52 |
36 |
42 |
50 |
12 |
38.4 |
41.6 |
||
|
Doubles boucles |
20 |
4 |
6 |
4 |
2 |
7.2 |
14 |
4 |
4 |
2 |
2 |
5.2 |
6.2 |
||
|
Tourbillons |
48 |
46 |
30 |
46 |
16 |
37.6 |
38 |
32 |
38 |
48 |
10 |
33.2 |
35.4 |
||
|
P.I. I. |
1.66 |
1.44 |
1.28 |
1.12 |
1.48 |
6.98 |
1.48 |
1.3 |
1.34 |
1.48 |
1.08 |
6.68 |
13.66 |
||
|
ES |
0.07 |
0.09 |
0.09 |
0.07 |
0.08 |
0.35 |
0.08 |
0.08 |
0.09 |
0.08 |
0.06 |
0.34 |
0.69 |
||
|
Ecart-type |
0.52 |
0.61 |
0.61 |
0.48 |
0.58 |
2.47 |
0.58 |
0.58 |
0.63 |
0.54 |
0.40 |
2.44 |
4.88 |
||
|
K-S |
3.00 |
2.26 |
2.25 |
2.96 |
2.37 |
1.13 |
2.37 |
2.39 |
2.03 |
2.34 |
3.25 |
1.66 |
1.32 |
||
|
|
*** |
*** |
*** |
*** |
*** |
NS |
*** |
*** |
** |
*** |
*** |
NS |
NS |
||
|
Femmes |
|||||||||||||||
|
|
DI |
DII |
DIII |
DIV |
DV |
D |
GI |
GII |
GIII |
GIV |
GV |
G |
D+G |
||
|
Total arcs |
6 |
20 |
12 |
4 |
10 |
10 |
2 |
22 |
10 |
4 |
4 |
8.4 |
9.2 |
|
||
|
Arcs plats |
6 |
18 |
12 |
2 |
8 |
8.8 |
2 |
20 |
10 |
4 |
4 |
8 |
8.4 |
|
||
|
Arcs en tente |
0 |
2 |
0 |
2 |
2 |
1.2 |
0 |
2 |
0 |
0 |
0 |
4 |
2.6 |
|
||
|
Boucles ulnaires |
34 |
46 |
76 |
42 |
72 |
54 |
26 |
32 |
54 |
40 |
80 |
46.4 |
50.2 |
|
||
|
Boucles radiales |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
4 |
4 |
2 |
0 |
2 |
1 |
|
||
|
Total tourbillons |
60 |
34 |
12 |
54 |
18 |
36 |
72 |
42 |
32 |
54 |
16 |
43.2 |
38.6 |
|
||
|
Doubles boucles |
16 |
8 |
2 |
4 |
0 |
6 |
34 |
12 |
4 |
0 |
2 |
10.8 |
8.4 |
|
||
|
Tourbillons |
44 |
26 |
10 |
50 |
18 |
30 |
38 |
30 |
28 |
54 |
14 |
32.4 |
31.2 |
|
||
|
P.I. I. |
1.54 |
1.16 |
1 |
1.52 |
1.1 |
6.32 |
1.7 |
1.22 |
1.22 |
1.5 |
1.12 |
6.76 |
13.08 |
||
|
ES |
0.09 |
0.10 |
0.07 |
0.08 |
0.07 |
0.35 |
0.07 |
0.11 |
0.09 |
0.08 |
0.06 |
0.36 |
0.7 |
||
|
Ecart-type |
0.61 |
0.71 |
0.49 |
0.54 |
0.51 |
2.49 |
0..51 |
0.76 |
0.62 |
0.58 |
0.44 |
2.15 |
4.96 |
||
|
K-S |
2.64 |
1.76 |
2.69 |
2.48 |
2.82 |
1.04 |
3.14 |
1.88 |
2.26 |
2.44 |
3.17 |
0.95 |
0.80 |
||
|
|
*** |
* |
*** |
*** |
*** |
NS |
*** |
* |
*** |
*** |
*** |
NS |
NS |
||
Tableau 3. Fréquences des figures digitales (%) et indice de complexité (P.I.I.) par doigt et par sexe dans la population d’Asni . D = droit, G = gauche, Nombres I, II,III, IV et V sont relatifs au doigts, P.I.I. = Pattern Intensity Index (indice de complexité), SE = Erreur Standard, K-S = test de Kolmogrov-Smirnov , NS : test non significatif ; * : test significatif à 5% ; ** : test significatif à 1%, *** : test significatif à 1‰.
Concernant l’indice de complexité, par ordre de complexité décroissant,
on trouve chez les hommes les doigts I, II, V, III et IV pour la main droite et
I, IV, III, II et V pour la main
gauche. Chez les femmes, on trouve les doigts I, IV, II, V et III pour la main
droite et I, IV, II, III et V pour la main gauche. Lorsque
chacun des doigts est pris séparément, un écart significatif du nombre de
triradii par rapport à la distribution normale est noté chez les deux sexes
(test de KS). Lorsqu’ils sont considérés ensemble, aussi bien pour la main
droite, la main gauche que les deux mains, on note un alignement à la
distribution normale.
I- 2/ Fréquence des groupes sanguins dans la population d’Asni
Le tableau 4 regroupe les résultats pour les trois systèmes
de groupes sanguins ABO, Rhésus et Kell. Pour le système ABO, les
fréquences obtenues des 4 phénotypes sont de 37.2%, 19.5%, 2.7% et 40.7%
respectivement pour les groupes A, B, AB et O. Les fréquences des allèles sont
donc de 63% pour ABO*O, 24% pour ABO*A et 13% pour ABO*B. Pour le système
Rhésus, 88,4% d’individus sont Rh+ et 11,6% sont Rh-. Les fréquences pour les 3
gènes C, D et E sont respectivement de 0.393 et 0.607 pour les allèles C et c, de
0.659 et 0.341 pour les allèles D et d et de 0.112 et 0.888 pour les allèles E et e. Pour le système Kell, les fréquences des phénotypes
K+ et K- sont respectivement de 94.6% et 5.4%. Les fréquences des allèles sont de 97% pour K+ et de 3% pour K-.
Les tests du ![]() (équilibre génétique de Hardy Weinberg) montrent que les deux systèmes
ABO et Kell sont panmictiques, alors que le système Rhésus est en déséquilibre.
(équilibre génétique de Hardy Weinberg) montrent que les deux systèmes
ABO et Kell sont panmictiques, alors que le système Rhésus est en déséquilibre.
|
Systèmes |
Phénotypes |
Effectifs observés |
Effectifs théoriques |
Fréquences alléliques |
|
ABO |
A |
42 |
40.680 |
ABO*A = 0.24 |
|
B |
22 |
20.419 |
ABO*B = 0.13 |
|
|
AB |
3 |
7.050 |
ABO*O = 0.63 |
|
|
O |
46 |
44.849 |
|
|
|
Total |
113 |
112.999 |
|
|
Rhésus
|
DCC-EE |
0 |
0.326 |
Rh*D = 0.6594 |
|
DCC-Ee |
0 |
2.188 |
Rh*d = 0.3406 |
|
|
DCC-ee |
20 |
17.577 |
|
|
|
DCc-EE |
0 |
0.429 |
Rh*C = 0.3929 |
|
|
DCc-Ee |
6 |
3.412 |
Rh*c = 0.6071 |
|
|
DCc-ee |
42 |
27.160 |
|
|
|
Dcc-EE |
1 |
0.662 |
Rh*E = 0.1119 |
|
|
Dcc-Ee |
17 |
5.272 |
Rh*e = 0.8881 |
|
|
Dcc-ee |
13 |
41.967 |
|
|
|
CCdd-EE |
0 |
0.036 |
|
|
|
CCdd-Ee |
0 |
0.290 |
|
|
|
CCdd-ee |
0 |
2.307 |
|
|
|
Ccdd-EE |
0 |
0.056 |
|
|
|
Ccdd-Ee |
0 |
0.448 |
|
|
|
Ccdd-ee |
0 |
3.564 |
|
|
|
ccdd-EE |
0 |
0.087 |
|
|
|
ccdd-Ee |
0 |
0.692 |
|
|
|
ddcc-ee |
13 |
5.507 |
|
|
|
Total |
112 |
111.980 |
|
|
|
Kell |
K- |
105 |
105.450 |
Kell*K- = 0.97 |
|
K+ |
6 |
6.429 |
Kell*K+ = 0.03 |
|
|
Total |
111 |
111.879 |
|
Tableau 4. Fréquences alléliques des systèmes sanguins (ABO, Rhésus et Kell) dans la population d’Asni. NS : test non significatif ; *** : test significatif à 1‰.
II/ Position de la
population d’Asni par rapport à diverses populations méditerranéennes
suivant les marqueurs étudiées (dermatoglyphes et les
systèmes sanguins ABO et Rhésus)
II-1/
Etude des figures digitales
Les fréquences des quatre types de dermatoglyphes ont été comparées à celles des
différentes populations de la base de données (figure 1). La classification
hiérarchique (dendrogramme) obtenue à partir de la matrice des distances met en
évidence deux grands groupes: un premier regroupant
les populations de l’Afrique du Nord (y compris la population étudiée), et un
deuxième comprenant les populations du sud de l'Europe. A l’exception des
populations marocaines qui présentent une grande diversité, on remarque
donc une grande cohérence géographique dans ces regroupements. La population d’Asni, bien que présentant des caractéristiques
propres qui la différencient des autres populations, se rattache plus au groupe
des populations du Sud de la Méditerranée (arabes et berbères) qu'au groupe des
populations du Nord (Espagne, France et Italie).
. 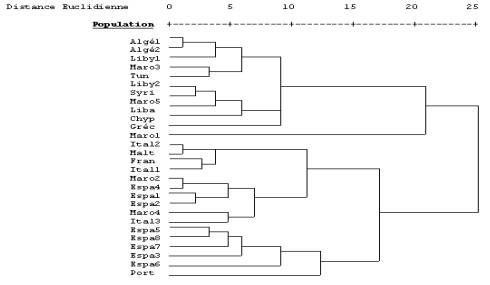
Figure 1. Dendrogramme (distances euclidiennes) obtenu à partir de la matrice
des fréquences des dermatoglyphes de quelques populations d’Afrique du Nord,
Moyen Orient et du Sud de l’Europe
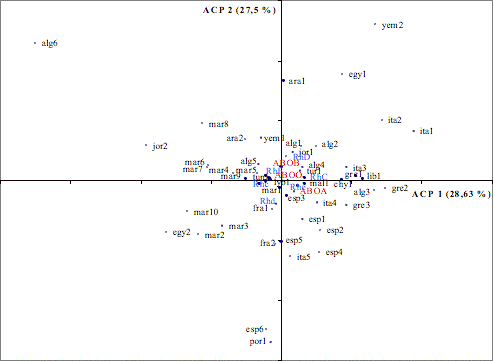
Figure 2. Plan factoriel engendré par les deux premiers axes de l’Analyse en
Composante Principale calculée à partir des fréquences des allèles des groupes
sanguins ABO et Rhésus.
II-2/ Etude des systèmes sanguins ABO et Rhésus
L’analyse en composante principale (ACP) a été réalisée pour situer
notre population dans l’ensemble Nord-Africain
et Méditerranéen à partir des fréquences des deux systèmes ABO et Rhésus. La
figure 2 donne la projection des populations dans le plan factoriel engendré
par les deux axes ACP1 et ACP2 contribuant simultanément pour 28,2% et 27,5% de
la variabilité totale.
L’axe ACP1 oppose les populations suivant les différences de fréquences
des gènes C/c et E/e du système Rhésus et de l’allèle ABO*A. Les allèles RhC et Rhe sont corrélés
positivement avec l’axe ACP1. Il oppose l’ensemble des populations
marocaines à certaines populations
du Nord de l’Afrique, du Moyen Orient et la majorité des populations du Sud de l’Europe.
L’axe ACP2 oppose les populations suivant les fréquences des allèles D et d du système Rhésus et la fréquence de l’allèle ABO*B.
Les allèles RhD et ABO*B sont corrélés positivement avec cet axe. Ce dernier
différencie d’une part, les populations marocaines entre elles, et d’autre
part, les populations du sud de l’Europe (surtout l’Italie par rapport à la France et l’Espagne).
La population d’Asni (mar1) ne se différencie pas suivant ces deux
axes. Elle se situe au milieu de l’ensemble des populations, indiquant des
valeurs moyennes pour les fréquences des allèles RhD, Rhd, RhC, Rhc et ABO*A
par rapport à l’ensemble de la base de données. L’analyse
en composante principale basée sur les fréquences des groupes sanguins confirme
donc la position intermédiaire de notre population, entre les populations
arabes et du Moyen-Orient et les populations du Sud de l’Europe.
Discussion et conclusion
Bien que les
dermatoglyphes et les groupes
sanguins ne reflètent qu’une faible
partie de la diversité biologique des populations humaines, nous pouvons
toutefois émettre quelques hypothèses à partir de la structure génétique des
diverses populations et de leurs liens de parenté. La comparaison des
populations du pourtour méditerranéen de la base de données montre tout d’abord
une différenciation des populations en deux grands groupes. Ils reflètent en
majorité les populations des deux rives de la Mer Méditerranée. Les résultats
sont similaires aussi bien pour l’analyse des dermatoglyphes que pour celle des groupes sanguins.
Cette différenciation génétique peut s’expliquer par l’existence d’une barrière
géographique à la migration : la Méditerranée aurait pu agir comme une barrière
géographique durant une période très éloignée de la notre. Elle aurait conduit
à une évolution indépendante des
populations après leur implantation (Bosch et al., 1997, Simoni et al., 1999). Cette barrière n’est évidemment pas
totale ; la structure
génétique des populations marocaines apparaît intermédiaire entre celle des
Européens du sud et celle des Nord Africains et des Moyen Orientaux. Il est
clair que les populations marocaines sont placées à un carrefour d’influences
provenant des populations du Proche-Orient, de l’Europe et de l’Afrique
sub-Saharienne. Tout au long de l’histoire, des grands flux migratoires
nord-sud et est-ouest ont existé. Arnaiz Villena et Al. (1999 a,b) rapportent
que le berbère se parlait des Iles Canaries (Guanche) jusqu’à l’Egypte, et de
la côte sud de la Méditerranéenne jusqu’à l’aire subsaharienne. Les peuples
berbères ont été forcés à émigrer vers 6 000 ans avant J.-C., lorsque les
conditions hyper arides du Sahara se sont établies. Ils se sont dirigés vers
les Iles Canaries, vers le Proche Orient, vers la péninsule ibérique et vers
les îles méditerranéennes. Les populations d’Afrique du Nord pourraient aussi
avoir peuplé certaines régions de l’Europe du Sud et traversé le détroit de
Gibraltar à une époque où il n’était pas encore sous les eaux (Chabaani et Cox,
1988). Une partie du patrimoine génétique et culturel des ibères est due aux
Berbères. La langue basque est très apparentée avec le berbère (Arnaiz Villena,
1999). La différenciation génétique de la population marocaine peut aussi avoir
une origine plus récente et être expliquée en partie par la conjonction de
nombreux autres facteurs, telles que les contraintes géographiques et
orographiques, ou encore l’hétérogénéité culturelle.
La proximité
génétique des populations arabophones et berbérophones est aussi un résultat
important à souligner, en accord avec la majorité des observations effectuées à
l’aide de nombreux autres marqueurs génétiques (Dugoujon
et al., 2003). Cette proximité indique que contrairement à l’idée reçue
d’une islamisation principalement culturelle du Nord de l’Afrique, celle-ci
aurait eu aussi un impact
génétique important.
En ce qui concerne la population d’Asni, bien qu’elle se situe
clairement au sein de la variabilité de la majorité des populations sud
méditerranéennes, celle-ci présente des fréquences qui la différencie des
autres populations. Cette différenciation peut s’expliquer par son isolement relatif dû aux conditions de vie particulières des
zones de haute montagne, propices à l’endogamie, aux effets fondateurs et à la
dérive génétique.
Il est également important
de souligner que l’histoire de quelques gènes ne traduit pas forcement celle
d’une population. D’où la nécessité de recourir à un grand nombre de gènes
variables pour retracer les migrations et mesurer les distances génétiques.
Références
bibliographiques
Arnaiz-Villena A, Martinez-Laso J,
Alonso-Garcia J. 1999a. Iberia: population genetics, anthropology, and
linguistics. Hum Biol. Oct;71(5):725-43.
Arnaiz-Villena A. 1999b. Berbère et génétique
historique. Traduit par Rachid Raha (Président du CMA). EL PAíS, 29-12.
Arrieta, M. 1985. Anàlisis dermatoglifico de la poblacion Vasca.
Tesis Doctoral. Universidad del Pais Vasco (Bilbao). España.
Auzas, C. 1957. Les Flittas, Etude
Ethnologique et Sérologique. Bull et Mem de la Soc d’Ant de Paris, T 8, 10eme
série, pp : 329-340.
Bonné, B., Ashbel,
S., Modal, M., Godber, M. J., Mourant, A. E., Tillis, D. and Woodhead, G. 1970.
Te Habbanite Isolate, Genetic Markers in Te Blood. Human Heredity, 20,
pp :609-622
Bosch, E., Calafell, F., Perez-Lezaun, A.,
Comas, D., Mateu E. et Bertranpetit J. 1997. Population history of North
Africa : Evidence from classical genetic markers. Hum. Biol., 69, pp :
295-311.
Bozicevic, D., Milicic,
J., N’dhlowu, J., Pavicic, D., Rudan, P. et Vassalo, A. 1993. Dermatoglyphic traits in the Malta population.
Coll. Antropol., 17 (1), pp : 137-146.
Camps, G., 1980. Les Berbères. Mémoire et identité. (ed).
Paris, 260p.
Chaabani H. et Cox D.W. 1988. Genetic characterization and
origin of Tunisian Berbers. Hum Hered
38: 308-316.
Chamla, M.CI. 1961.
Les empreintes digitales de 2336
Algériens musulmans. L’anthropologie, 65, pp : 444-466.
Chamla, M.CI.1973. Structure anthropologique
des Algériens du Nord. L’anthropologie, 77, : pp :717-754.
Colino, F. 1978. Antigenos eritrocitarios de
los grupos sanguineos en la poblacion Española. Tesis Doctoral, Univ,
Complutense, Madrid.
Cummins, H. et Midlo, C. 1961. Finger prints, palms and
soles. 2ème édition, New York, Dover Publications.
Cunha, X et Abreu, M.D. 1954. Impresoes
digitais de Portugueses. Pourcentagens de figuras, valores quantitativos e
frequencias empiricas dos genes VR e U. Comtribuçoes para Estudo da
Antropologia Portuguesa, 5 ,pp : 315-347.
Dugoujon J.M., Lemaire O., Guitard E., Sevin A., Larrouy G., Baali A., Sabir
B. et Cherkaoui M 2003. Etude de
la diversité des haplotypes des Immunoglobulines (systèmes Gm et Km) d’une population berbère de la vallée
de Tacheddirt (Haut Atlas, Maroc). Comparaison à d’autres populations Berbères,
Africaines et Européennes (soumis aux Bulletins et Mémoires de la Société
d’Anthropologie de Paris)
Egocheaga, J.E. 1973. Las lineas
dermopapilares en Asturianos. I. Dermatoglifos dactilares. Trab. Antropologia,
17, pp : 27-50.
Errahaoui, MY I., 2002. Caracterisation
anthropogénétiques de la population Berbères de Ouarzazate : Analyse
comparative du polymorphisme des dermatoglyphes et des groupes sanguins :
ABO, Rh, MNSs, et Duffy à l’échelle méditerranéenne. Mémoire de diplôme des
études supérieures approfondies. Université Chouaib Doukkali, Faculté des
Sciences ELJadida, Maroc.
Falco, G. 1917. Sulle figure papillare dei
popatrelli dell dita nei Libici. Revista Anthropologia, 22 : 91-148.
Fernandez-Santander, A., Kandil, M., Luna, F.,
Estiban, E., Gimenez, F., Zaoui, D. Et Moral, P. 1999. Genetic realationships between
southeastern Spain and Morocco: New data on ABO , Rh, MNSs, and DUFFY
polymorphisms. Am. J. Biol., 11, pp : 745-752.
Fuster, V. et Cabello, M.J. 1985. Deramatoglifos digitales en una
poblacion de la Sierra de Gredos. Actas IV Cong. Esp. Anthro. Biol.,
(Barcelona, España).
Gessain, M. 1957. Les crêtes papillaires
digitales de 194 marocains. Bull. et Mém. De la Soc. Anthropo. De Paris, 7,
pp : 262-271.
Gessain, M. et Gessain, R. 1956. Les crêtes
digitales et palmaires de 346 français. Bulletin et mémoires de la Société
d’Anthropologie de Paris, 8, pp : 341-353.
Gualdi-Russo, E., Zannotti, M. et Cenni S.
1982. Digital
dermatoglyphics in Italians. Hum. Biol., 54 (2), pp : 373-386.
Harich, N., Esteban, E.,
Chafik, A., Lopez-Alomar, A., Vona, G., Moral, P. 2002. Classical
polymorphisms in Berbers from Moyen Atlas (Morocco): genetics, geography and
historical evidence in the Mediterranean peoples. Ann Hum Biol. 29, pp: 473-487.
Johnson, R.H.,
Ikin, E, W., and Mourant, A. E.,1963. Blood Groups of the Ait Haddidu Berbères
of Morocco. Hum Biol, 35 (4), pp: 514-523.
Kandil, M. 1999. Etude anthropo-génétique de
la population arabe du Maroc
méridional (Abda, Chaouia, Doukkala et Tadla). Thèse d’Etat, Université Chouaib
Doukkali, Faculté des Sciences ELJadida., Maroc.
Kandil, M., Luna, F., Chafik, A., Zaoui, D. et
Moral, P. 1998. Digital dermatoglyphic patterns of Morrocan Arabs : relationships
with Mediterranean populations. Annals of Human Biology, 25 (4), pp :
319-329.
Luna, F. et Pons, J. 1987. The dermatoglyphics of the Eastern
Andalusia. International J. of
Anthro., II (2), pp : 183-190.
Manzano, C., Aguirre, A. I., Iriondo, M.,
Martin, M., Osaba, L. et De La Rua, C. 1996. Genetic polymorphisms of
Basques from Gipuzkoa : genetic heterogeneity of the Basque population. Ann. Hum. Biol., 23 (4),
pp : 285-296.
Mechali, MM. D., Leveque , J. Et Faure P.
1957. Les groupes sanguins ABO et
Rh des Haratin du Maroc. Bull et Mem de la Soc d’Ant de Paris., 196-204.
Memmi, M. 1999. Etude génétiquede la
population Corse et sa relation avec les populations Euro-méditerrannéennes.
Thése, Ph. D., Université de Corse, France.
Moral P. 1986. Estudio
antropogenetico de diversos polimorfismos hematologicos en la isla de Manorca.
Tesis Doctoral, Universidad de Barcelona . España .
Moral, P., Marogna, G., Salis, M.,
Succa, V. et Vona G. 1994. Genetic data on Alghero population
(Sardinia) : Contrast between biological and cultural evidence. Am. J. Phys. Anthropol., 7, pp : 25-33.
Moreno, P. et Moral P. 1983. Distribucion de
grupos sanguineos y enzimas eritrocitarios en una poblacion de Gerona (España).
Sep Garcia de Orta, Ser, Antropobiol., Lisboa, 2 (1-2), pp : 275-282.
Moreno, P. et Pons, J. 1985. Dermatoglifos
digitales y palmares en habitantes de Menorca. Act. IV Cong. Esp. Anthro.
Biol., (Barcelona, España ).
Mourant, A. E., Kopek, A. C. et Domaniewska-Sobkzak, K. 1976. Te distribution of
te humain Blood groups and other polymorphisms. Oxford Univ. Press. London.
Nabulsi, A., Cleve, H. et
Rodewald, A. 1997. Serological analysis of te Abbad tribe of Jordan. Hum. Biol.,
69, pp : 357-373.
Naffah, J. 1974.
Dermatoglyphics and flexion creases in the Lebanese population. Am. J. phys. Anthrop., 41, pp : 391-410.
Oyhenart, M.F. 1983. Dermatoglifos en
Gallegos. I. Caracters de apreciacion cualitativa. Actas III Cong. Antro.
Biol., (Santiago de Compostela, España).
Oyhenart, M.F.1985. Aproximacion a la
descripcion dermatoglifica de los Españoles del SudOeste : Extremeños y
Andaluces béticos. Tesis Doctoral, Fac. Biologia, Univ, de Barcelona, Eapaña.
Pascon, P. 1977. Le Haouz de Marrakech, Ed.
Marocaines et international, Rabat. 2 Tomes.
Penrose, L.S. 1968.
Memorandum on dermatoglyphic nomenclature. Birth defects, Original Article.
Series IV/3, pp : 1-13.
Piazza, A.,
Olivetti, E., Barbanti, M., Reali, G., Domenici, R., Berciolini, P., Caenazzo,
L., Corvito, P., Bestitti, A., Bonavita, V., Crino, C., Pascali, V., Fiori, A.
Et Bargagna, M.1989. The distrrbution of some polymorphisms in Italy. Gene
Geography, 3 : 69-139.
Plato, C. C. 1970.
Dermatoglyphics and flexion creases of Cypriot. Am. J. Phys. Anthropol., 23,
pp : 421-427.
Pons, J. 1952. Impresiones dermopapilaires en estudiantes universitarios barceloneses.
Trab. Instit. Bernardino de Sahagun, 13 : 87-129.
Portabales, M.D. 1983. Dermatoglifos en la
poblacion de La Alcarria (Guadalajara) : Configuraciones digitales. Bol.
de la Soc. Esp. de antropologia Biologica, 4 : 41-50.
Poumpouridou A. Et
Scheil H. G. 1995. the distribution of the ABO and Rhesus blood groups
(phenotype and allele frequencies) in the population of Cyprus. Gene Geography, 9,
pp : 197-205.
Roychoudhury,
A. K. et Nei, M. 1988. Human polymorphic genes world distribution. New York.
Oxford University Press.
Saha N, Bayoumi RA, El Sheikh FS, Samuel AP,
El Fadil I, El Houri IS, Sebai ZA, Sabaa HM. 1980. Some blood genetic markers
of selected tribes in Western Saudi Arabia. Am J Phys Anthropol. May;
52(4):595-600.
Salis, N., Floris, G., Boetsch, G., Rabino
Massa, E. 2001. Position d’une population Alpine par rapport à l’étude des
dermatoglyphes. XXVe
Colloque du Galf, Marseille (Francia) 16-18 Luglio : 79.
Shanklin, W.M. et Cummins, H. 1937.
Dermatoglyphics in Rwala bedouins. Human Biology, 9 (3), pp : 357-365.
Simoni, L.,
Gueresi, P., Pettener, D., and Barbujani G. 1999. Patterns of gene flow
inferred from genetic distances in the Mediterranean region. Human Bioloy, V :
71, N° : 3, pp : 399 - 415.
Solignac M et al.,1995. Génétique et évolution.
Hermann.
Tills, D., Kopec,
A.C. et Tills, R. E. 1983. Te distribution of human blood groups and other
polymorphisms. Supplement 1. Oxford Univ. Press. Oxford.
Tsiakalos G, Walter H, Hilling M, Windhof O.
1980. Investigations on the distribution of genetic polymorphisms in Greece. 1.
Blood group polymorphisms. Anthropol Anz. Nov; 38(4):237-50.
Vona,
G., Calo, C.M., Autuori, L., Mameli, G.E., Lixi, M.F., Ghiani, M.E., et Di Gaetano. C. 1998. Genetic
structure of western Sicily. International Journal of Anthropology ; 13
(2) : 137-147.
Vrydagh-Laoureux, S. 1979. Digital and palmar
dermatoglyphics: a sample of moroccans. Human Biology, 4 (51), pp :
537-549.