
Colloques du Groupement des Anthropologistes de Langue
Française (GALF)
Sardi
Marina, L., Ramírez-Rozzi, F., Pucciarelli Héctor, M.,
2004, Morphologie crânio-faciale des populations de l'Europe et du
Maghreb depuis le Paléolithique Supérieur jusqu'à présent. Antropo, 7, 145-153. www.didac.ehu.es/antropo
Morphologie
crânio-faciale des populations de l'Europe et du Maghreb depuis le
Paléolithique Supérieur jusqu'à présent
Sardi
Marina, L.1,2, Ramírez-Rozzi, F.1, Pucciarelli Héctor, M.2
1
UPR 2147 “Dynamique des Populations Humaines”, Centre National de la Recherche
Scientifique. 44, rue de l’Amiral Mouchez, 75014, Paris, France.
2
Departamento Científico de Antropología, Museo de La Plata, Facultad de Ciencias
Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque S/N, 1900
La Plata, Argentina.
Adresser
correspondance à:
msardi@fcnym.unlp.edu.ar
Mots
clés:
Paléolithique Supérieur, Néolithique, composante crânienne fonctionnelle,
morphologie.
Key
words: Upper
Paleolithic, Neolithic, Functional Cranial Components, morphology.
Résumé
Il n’y a pas de consensus sur les processus responsables de la Néolithisation en Europe et en Afrique du Nord. Le passage Paléolithique Supérieur-Néolithique a pu s’opérer par une migration des peuples du Proche-Orient ou par des changements économiques des populations locales. Nous avons effectué l’analyse de la taille et de la conformation crâniennes des populations qui ont habité ces deux régions depuis le Paléolithique Supérieur jusqu’à présent. Notre travail se fonde sur l ‘étude des composantes crâniennes fonctionnelles (CCF) suivantes: neurale antérieure, moyenne et postérieure, otique, optique, respiratoire, masticatoire et alvéolaire. Les différences observées ne mettent en évidence ni des tendances temporelles ni régionales. On observe, par contre, une nette différence entre les groupes du Paléolithique Supérieur et les autres. Les premiers se caractérisent par une taille plus grande et leur conformation est influencée par des CCF masticatoire et neurale moyenne importantes et une CCF optique réduite. Le développement de la CCF masticatoire est associé à une large face chez les groupes du Paléolithique Supérieur. Notre étude ne permet pas de résoudre la discussion sur la Néolithisation. Néanmoins, elle permet de suggérer que les différences dans la morphologie résultent d’un nouveau type de vie (nutrition, mobilité, contraintes masticatoires) à partir du Néolithique.
Abstract
It is not clear if the Neolithic transition in Europe and
North Africa resulted by a migration of peoples from the Near East, or by
economic changes of local populations, or by an interaction of both processes.
Size and shape variation of the Functional Cranial Components (FCC) of the
neurocranium and the face, from the Upper Paleolithic to modern periods in
these two regions, was assessed. The skull was divided in the anteroneural,
midneural, posteroneural, otic, optic, respiratory, masticatory and alveolar
FCCs. The differentiation does not express any temporal or regional pattern.
But a shift is observed between the Upper Palaeolithic groups and the later
samples. The former are characterised by greater size and the shape is mainly
affected by a greater midneural and masticatory FCCs, and smaller optic FCC.
The greater masticatory volume is associated to wider faces in the Upper
Palaeolithic groups. Our study cannot enable to conclude if the morphological
shift is caused by a replacement or by a change in the local populations, however
the morphological changes can be attributed to modifications in nutrition,
mobility and masticatory stress from the Neolithic period.
Le Paléolithique Supérieur et le Mésolithique européens sont caractérisés par une importante variabilité des fossiles du point de vue morphologique (Riquet, 1970; Lahr, 1996). La transition au Néolithique est principalement considérée comme le résultat d’une migration des peuples agriculteurs arrivés du Proche Orient (Price, 2000; Bentley et al., 2002). Nonobstant, même si les fréquences génétiques mettent en évidence la diffusion démographique (Ammerman et Cavalli Sforza, 1984), la contribution des populations Paléolithiques à la variabilité moderne ne pourrait être exclue (Chikhi et al., 1998; De Benedetto et al., 2000; Semino et al., 2000; Simoni et al., 2000). En fait, il y a des études qui montrent que le remplacement des populations Mésolithiques n’a pas eu lieu dans quelques régions de l’Europe, donc l’agriculture et/ou la céramique auraient été adoptées localement sans remplacement de populations (Jackes et al., 1997; Price, 2000; Zvelebil et Lillie, 2000; King et Underhill, 2002).
Le Paléolithique Supérieur du Maghreb est représenté par les Ibéromaurusiens. A la fin du Pléistocène tardif quelques vestiges sont attribués à de nouveaux chasseurs: les Capsiens. Venu du Proche Orient, ils se caractérisent par une morphologie plus gracile que celle des Ibéromaurusiens (Ferembach, 1986). Néanmoins, d’autres chercheurs suggèrent que les Ibéromaurusiens et les Capsiens faisaient partie de la même population (Sheppard et Lubell, 1990; Irish, 2000). La production alimentaire est reconnue chez les Capsiens tardifs, environ 6000 ans AP (Lubell et al., 1992) ou même avant, la subsistance étant basée sur le pastoralisme (Roubet, 2001).
Pendant l’Holocène initial, une tendance à la brachycéphalie
a été suggérée en Europe et en Afrique du Nord comme conséquence de la
gracilisation squelettique due à son tour à la diminution de l’activité
masticatoire (Riquet 1970, Lahr 1996).
En ce qui concerne l’étude de la morphologie crânienne, un
nouveau paradigme a été proposé par Moss avec l’hypothèse de la Matrice
Fonctionnelle (Moss, 1973; 1997 ; Moss et Young, 1960). Moss a suggéré que
la conformation du crâne résulte des fonctions de support et de protection que
les différentes parties du crâne doivent accomplir en regard des tissus mous et
des cavités. Ainsi, chaque fonction (mastication, vision, etc.) est effectuée
par ce que Moss (1973) a appelé une composante crânienne fonctionnelle (CCF)
qui comprend la matrice fonctionnelle (cavités et tissus qui exécutent une
fonction) et l'unité squelettique (tissus durs qui donnent la protection
biomécanique et supportent la matrice fonctionnelle). Cette hypothèse considère
que la croissance de l'os et du cartilage n'est pas exclusivement régi par
leurs propres facteurs génétiques. Les études expérimentales et cliniques
indiquent que la régulation de la croissance squelettique, tout au long de
l’ontogénie, suit les contraintes établies par la matrice
fonctionnelle.
L’objectif de cette étude, basée sur l’hypothèse de la
Matrice Fonctionnelle, est de déterminer s’il y a eu un changement important de
la morphologie crânio-faciale des populations de l’Europe et du Maghreb,
depuis le Paléolithique Supérieur jusqu’à présent. L’analyse de CCF devrait
permettre, à la différence des méthodes classiques, une compréhension plus
précise des types de changement qui ont affectés la morphologie crânienne des
populations humaines.
L’étude comprend des individus du Paléolithique Supérieur
européen
(Cro-Magnon 1, Solutré 5, Chancelade, Laugerie-Basse 2,
Saint-Germain-La-Rivière, Predmosti 3 et Mladec 1) (PSE) (n = 7); du Mésolithique (Teviec-Hoëdic) (TH) (n =
13); du Néolithique de la Lozère (LZR) (n = 25) (Homme Mort Cave et
Baumes Chaudes) et de la Vallée de la Marne (MRM) (n = 97); et des français
modernes des
XVIIème et XIXème siècles (n = 50).
Les échantillons d’Afrique du Nord sont : les Iberomaurusiens d’Afalou (AFL) (n = 27) et Taforalt (TAF) (n = 15); les Guanches
des Iles Canaries
(GU) (n = 51), population descendant des Berbères avec une économie basée sur
une agriculture élémentaire et la domestication du mouton et de la chèvre; les Puniques de Carthage (CTG) (n = 26), datés entre le
VIIIème et le IIIème siècles, et d’origine phénicienne; des individus de la
ville Al Qayrawan
(QRN) (n = 20), fondée par les Arabes dans l’année 670 A. D; et des Berbères
modernes de Kabylie
(KBL) (n = 20), Algérie.
Le crâne a été divisé en deux composantes majeures, neurale (N) et faciale (F). Chaque composante majeure a été divisée à son tour en quatre composantes mineures. La composante neurale comprend la neurale antérieure (NA), la neurale moyenne (NM), la neurale postérieure (NP) et la composante otique (OT). En ce qui concerne la composante majeur faciale, elle est formée par les composantes optique (O), respiratoire (R), masticatoire (M) et alvéolaire (A). Même si la division du CCF neurale ne présente pas un rapport bien définie avec la fonction, elle permet une meilleure compréhension de la spécificité et de la dynamique de la variation de chaque région crânienne. Chaque CCF a été mesurée par trois variables orthogonales qui déterminent la longueur, la hauteur, et la largeur.
Des indices volumétriques et morphométriques ont été obtenus
(Tableau 1). Les indices Volumétriques correspondent à la moyenne géométrique
des trois dimensions et mesurent donc la taille. Les indices Morphométriques donnent une indication de la
conformation des CCF majeures en termes de taille relative de ses quatre
composantes mineures. La conformation a été aussi estimée avec une
standardisation de la différence proportionnelle entre chaque dimension et
l’indice volumétrique (Tableau 1).
|
Indices |
calculs |
|
volumétrique |
(IV)3 (longueur x largeur x
hauteur) |
|
morphométriques neurales |
IV mineur / IV NA, NM, NP et OT |
|
morphométriques faciales |
IV mineur / IV O, R, M et A |
|
standardisation |
longueur / IV, largeur / IV, et hauteur
/ IV |
Tableau 1. Calculs
pour l’estimation de la taille absolue et relative et de la conformation des
CCFs.
CCF
(FCC) : composante cranienne-fonctionnelle. IV : indice volumétrique.
NA : neurale antérieure. NM : neurale moyenne. NP : neurale
postérieure. OT : otique. O : optique. R : respiratoire.
M : masticatoire. A : alvéolaire.
Table 1. Estimation
of the shape, absolute size, and relative size of FCCs.
La normalité des distributions de fréquences et
l’homogénéité des variances ont été vérifiées avec les analyses de
Kolmogorov-Smirnov et de Levene. La comparaison entre les populations a été
effectuée avec l’Analyse de la Variance. Une analyse Canonique a été effectuée
avec les indices volumétriques et les variables standardisées pour évaluer si
la variation uni-dimensionnelle des CCFs majeures est liée à la taille des CCFs
mineures.
Tous les indices montrent des différences significatives
(p < 0.01) pour le sexe masculin. Dans la Figure 1, les valeurs F et la
moyenne des indices volumétriques de chaque échantillon masculins sont
signalés. La taille neurale et la taille faciale diffèrent entre les
populations paléolithiques et post-paléolithiques, tandis qu’elles ne sont pas
trop différentes parmi les populations post-paléolithiques.
L’analyse des indices morphométriques neuraux suggère: 1.
l’indice neural moyen est le plus variable, il se distingue chez les
paléolithiques par sa taille supérieure et chez les Guanches, par sa taille
inférieure (Figure 2) ; 2. l’indice otique est plus petit chez les
individus du Paléolithique européen, du Néolithique de la Lozère et chez les
Kabyles ; 3. la composante neurale antérieure présente une taille relative
petite chez les individus de Taforalt et grande chez les Guanches; 4. la
composante neurale postérieure a une taille réduite chez les spécimens de
Taforalt et de la Kabylie et une grande taille chez ceux de la Lozère.
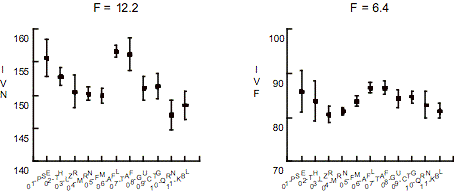
Figure 1. Valeurs F
de ANOVA, moyennes et limites de confiance (p = 0.95) pour les indices
volumétriques des CCFs majeures. IVN : indice volumétrique neurale.
IVF : indice volumétrique facial. PSE : Paléolithique supérieur européen.
TH : Mésolithiques de Teviec-Hoëdic. LZR: Néolithique de la
Lozère. MRN : Néolithique de la Marne. FM : Français modernes.
AFL : Paléolithiques d’Afalou. TAF : Paléolithiques de Taforalt.
GU : Guanches. CTG : Puniques de Carthage. QRN : arabes de Al Qayrawan.
KBL: berbères de Kabylie.
Figure 1. F values of
ANOVA, averages and confidence limits at p = .95 for volumetric indices of
major FCCs.
Par rapport aux indices morphométriques faciaux, l’indice
masticatoire montre la variation la plus large avec une taille considérable
chez les chasseurs-cueilleurs (paléolithiques et mésolithiques) (Figure 2). La
composante optique a une taille inférieure chez les paléolithiques. Les
individus d’Afalou et de Taforalt présentent des composantes alvéolaire et respiratoire
plus petites, respectivement.
La corrélation canonique réalisée avec les indices
volumétriques neuraux et les variables standardisées a permis d’obtenir 3 axes,
les deux premières étant significatives. Les coefficients de corrélation
canonique (R) sont pour le premier axe 0,49 (![]() = 117,7, p < 0,01), pour le deuxième axe 0,18 (
= 117,7, p < 0,01), pour le deuxième axe 0,18 (![]() = 16,1, p < 0,05) et pour le troisième axe 0,12 (
= 16,1, p < 0,05) et pour le troisième axe 0,12 (![]() = 4,9). Le Tableau 2 représente la contribution des variables
et des indices aux deux axes significatifs. La Figure 3 représente la première
variable canonique qui montre les paléolithiques européens, Kabyles et
néolithiques de la Marne séparés du reste. La conformation neurale de ces
groupes se caractérise par une composante neurale moyenne plus grande associée
à la hauteur neurale.
= 4,9). Le Tableau 2 représente la contribution des variables
et des indices aux deux axes significatifs. La Figure 3 représente la première
variable canonique qui montre les paléolithiques européens, Kabyles et
néolithiques de la Marne séparés du reste. La conformation neurale de ces
groupes se caractérise par une composante neurale moyenne plus grande associée
à la hauteur neurale.
La corrélation canonique réalisée avec les indices et les
variables faciales a permis d’obtenir deux axes significatives. Les
coefficients de corrélation canonique (R) sont pour le premier axe 0,42 (![]() = 92,7, p < 0,01), pour le deuxième axe 0,23 (
= 92,7, p < 0,01), pour le deuxième axe 0,23 (![]() = 22,6, p < 0,01) et pour le troisième axe 0,07 (
= 22,6, p < 0,01) et pour le troisième axe 0,07 (![]() = 1,8). La contribution des variables et des indice aux axes
significatifs (Tableau 2) indique que le volume masticatoire est corrélé de
façon directe avec la largeur faciale, et le volume optique est corrélé de
façon inverse avec la longueur faciale. Le premier axe sépare les
chasseurs-cueilleurs du reste des échantillons. Les chasseurs-cueilleurs
présentent une face plus large avec un volume masticatoire majeur (Figure 3).
= 1,8). La contribution des variables et des indice aux axes
significatifs (Tableau 2) indique que le volume masticatoire est corrélé de
façon directe avec la largeur faciale, et le volume optique est corrélé de
façon inverse avec la longueur faciale. Le premier axe sépare les
chasseurs-cueilleurs du reste des échantillons. Les chasseurs-cueilleurs
présentent une face plus large avec un volume masticatoire majeur (Figure 3).
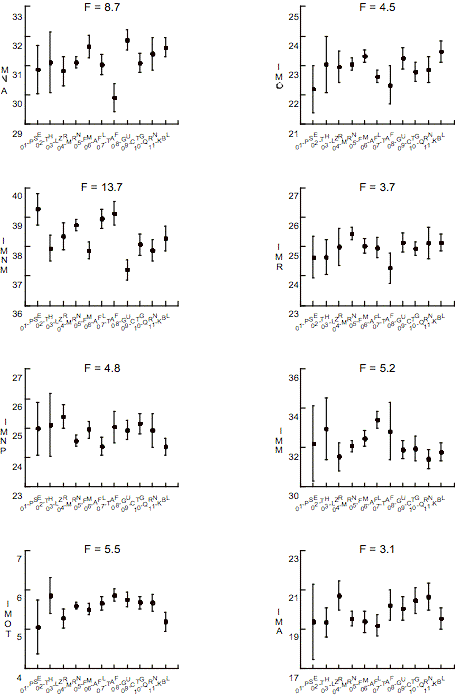
Figure 2. Valeurs F
de ANOVA, moyennes et limites de confiance (p = 0.95) pour les indices
morphométriques (IM). IMNA : IM neural antérieur. IMNM : IM neural
moyen. IMNP : IM neural postérieur. IMOT : IM otique. IMO : IM
optique. IMR : IM respiratoire. IMM : IM masticatoire. IMA : IM
alvéolaire. Voir Fig. 1 pour abréviations.
Figure 2. F values of
ANOVA, averages and confidence limits at p = .95 of morphometric indices. See
Fig. 1 for abbreviations.
|
|
CCF Neurale |
CCF Faciale |
||
|
|
Axes canoniques |
|||
|
|
1 |
2 |
1 |
2 |
|
IVNA |
0.19 |
0.12 |
|
|
|
IVO |
|
|
-0.08 |
0.84 |
|
IVNM |
-0.56 |
0.44 |
|
|
|
IVR |
|
|
-0.28 |
0.02 |
|
IVNP |
0.48 |
0.63 |
|
|
|
IVM |
|
|
0.47 |
0.31 |
|
IVOT |
-0.08 |
0.71 |
|
|
|
IVA |
|
|
-0.53 |
0.4 |
|
longueur neurale |
0.75 |
-0.39 |
|
|
|
longueur faciale |
|
|
0 |
-0.99 |
|
largeur neurale |
0.12 |
0.74 |
|
|
|
largeur faciale |
|
|
0.93 |
-0.33 |
|
hauteur neurale |
-0.91 |
-0.35 |
|
|
|
hauteur faciale |
|
|
-0.66 |
-0.74 |
Tableau 2.
Contribution aux axes significatifs de 2 groupes de variables comparées dans la
Corrélation Canonique. Voir Tableau 1 pour les abréviations.
Table 2. Loadings on
signicant canonical variates of two groups of variables. See Table 1 for
abbreviations.
La taille élevée des CCFs chez les paléolithiques est en
accord avec d’autres études (Smith, 1979; Lahr, 1996). Elle peut être associée
à des facteurs systémiques liés probablement à leur mode de vie. L’augmentation
volumétrique de la matrice fonctionnelle et/ou la déposition de tissu
squelettique peuvent causer la majeure taille crânienne (Moss et al., 1987).
Une alimentation insuffisante en protéines pourrait mener à une taille plus
petite parmi les groupes néolithiques et modernes. Mais pour connaître son
effet, il serait nécessaire d’évaluer la corrélation entre la taille et plusieurs
indicateurs de santé et morphologiques du squelette. L'action hormonale
correspond à un autre facteur systémique. En effet, il y a un lien direct entre
la circulation de l'hormone de croissance et l'activité physique, en
particulier la mobilité (Weltman et al., 2001). Chez les humains, l’hormone de
croissance, dont la circulation a été enregistrée depuis la naissance (Geary et
al., 2003), favorise l’augmentation de la masse squelettique (Vogl et al
1993 ; Lieberman, 1996 ; Barr et McKay, 1998 ; Banu et al.,
2001) et musculaire, et influence aussi la croissance craniofaciale (Vogl et
al, 1993). De ce fait, il faudrait s’attendre à que les fermiers présentent une
réduction plus marquée de la face que du crâne neural, car la face se
caractérise par une période de croissance plus longue et tardive. De toutes
façons, même si les facteurs alimentaires semblent plus probants pour
corroborer la réduction de la taille dans le post-mésolithique, l'effet des
facteurs hormonaux ne peut pas être mis à l’écart complètement.
La conformation crânio-faciale a été évaluée par les indices
morphométriques et par la standardisation des variables. Dans la conformation
de la composante majeure neurale, la composante neurale moyenne montre une
relation inverse avec les composantes neurales antérieure et postérieure. Des
nombreuses études proposent que la voûte montre des variations associées aux
changements de la taille et la conformation du cerveau et/ou de la base
crânienne (Moss et Young, 1960; Lieberman et al., 2000a; 2000b). Il est
probable que la composante neurale moyenne présente une capacité majeure à
s’adapter face aux changements des structures neurales et squelettiques
associées, tandis que les modifications des composantes antérieure et
postérieure seraient dues aux changements compensatoires de taille relative.
Depuis le XIXème siècle, des auteurs ont proposé que la dolichocéphalie caractérise les paléolithiques tandis que la brachycéphalie caractérise les néolithiques et les populations modernes (Broca, 1873; Riquet, 1970; Lahr, 1996). Différemment, nos résultats suggèrent que la hauteur plutôt que la longueur ou la largeur neurale correspond à l’aspect le plus important pour discriminer les populations paléolithiques de celles du Néolithique. En plus, la distribution ne révèle pas des tendances temporelles ou régionales pour ces caractéristiques.
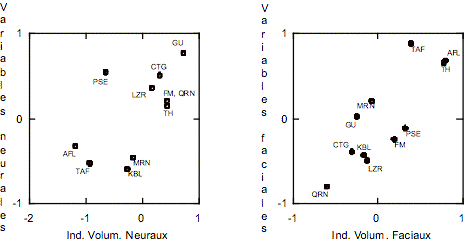
Figure 3.
Distribution des moyennes des échantillons dans le premier axe de l’analyse
canonique. Voir Figure 1 pour abréviations.
Figure 3. Plot of
average samples on the first canonical variate. See Figure 1 for abbreviations.
Une composante masticatoire importante distingue les chasseurs-cueilleurs des post-mésolithiques. Elle est accompagnée par une augmentation de la largeur faciale. La variation absolue et relative de taille de la FCC masticatoire chez les chasseurs-cueilleurs peut être attribuée aux différences dans le stress masticatoire. En effet, les forces masticatrices produisent des changements crânio-faciaux et leur intensification induisent l’augmentation de la taille des muscle et des structures apparentées. Selon Raadsheer et al. (1999), l’intensité des forces masticatrices est intimement corrélée avec l'épaisseur des muscles et les dimensions transversales et verticales de la face. Kiliaridis (1995) a proposé que l'hyperfonction masticatoire mène à un agrandissement de la face par croissance suturale et déposition de tissu squelettique. Les études expérimentales ont démontré qu'un régime alimentaire plus molle peut contribuer, parmi d’autres changements, à une réduction de l'épaisseur corticale de l’os et de la densité mandibulaire (Bresin et al., 1999), au serrage des arcs maxillaires et à la déformation du palais (Beecher et al, 1983), à la réduction de la taille musculaire (Ciochon et al, 1997), et à des modifications du condyle mandibulaire (Giesen et al., 2003). D'autres études où des populations avec des modes de subsistance différents ont été comparées, ont suggérée des différences dans les structures masticatoires (Carlson et Van Gerven, 1979; Sardi et al., article soumis). Donc, le mode de vie des chasseurs cueilleurs, avec un haut stress masticatoire, explique bien la morphologie décrite.
Une CCF optique de petit volume caractérise les individus paléolithiques et Kabyles. L’orbite est affectée par le lobe frontal qui participe de la composante neurale antérieure (Lieberman et al., 2000b). Celle-ci aussi présente une petite taille dans ces populations. La taille de la CCF optique est peut-être associée à l’origine neurale de sa matrice fonctionnelle.
Notre étude ne permet pas de suggérer la présence d’une morphologie particulière pour chaque région, ni de l’existence d’une tendance temporale. Néanmoins, une différence significative et importante a été obtenue entre les paléolithiques et les post-mésolithiques. Elle concerne la taille et la conformation crânienne. Ces différences peuvent être mises en relation avec les changements de la nutrition, de la mobilité et du stress masticatoire.
Hélas, nos résultats ne permettent pas de résoudre le débat sur la transition Néolithique. Les changements morphologiques observés pourraient être expliqués par un remplacement des populations. Mais il est également probable que la transition au Néolithique ait été effectuée par des populations locales, la grande différence morphologique étant le résultat des changements de leur mode de vie.
Remerciements. Nous
remercions à Philippe Mennecier, du Musée de L’Homme, et à Henry de
Lumley et Dominique Grimaud-Hervé, de l’Institut de Paléontologie Humaine, pour
faciliter l’accès aux collections. Cette étude a été possible grâce aux bourses
de la Fondation Fyssen et de l’Universidad Nacional de La Plata données
à MLS et à l’accord de coopération CONICET-CNRS.
Bibliographie
Ammerman, A. J., et Cavalli
Sforza, L. L., 1984, The Neolithic Transition and the Genetics of Population in
Europe (Princeton: Princeton University Press).
Banu, J., Orhii, P. B.,
Okafor, M. C., Wang, L., et Kalu, D. N., 2001, Analysis of the effects of
growth hormone, exercise and food restriction on cancellous bone in different
bone sites in middle-aged female rats. Mechanisms of Ageing and Development,
122, 849-864.
Barr, S. I., et McKay, H. A.,
1998, Nutrition, exercise, and bone status in youth. International Journal of
Sport Nutrition, 8, 124-142.
Beecher, R. M., Corrucini, R.
S., et Freeman, M., 1983, Craniofacial correlates of dietary consistency in a
nonhuman primate. Journal of Craniofacial Genetics and Developmental Biology,
3, 193-202.
Bentley, R. A., Price, T. D.,
Gronenborn, D., Wahl, J., et Fullagar, P. D., 2002, Prehistoric migration in
Europe: strontium isotope analysis of Early Neolithic skeletons. Current
Anthropology, 43, 799-804.
Bresin, A., Kiliaridis, S., et
Strid, K. G., 1999, Effect of masticatory function on the internal bone
structure in the mandible of the growing rat. European Journal of Oral
Sciences, 107,
35-44.
Broca, P., 1873, Les crânes de
la Caverne de l’Homme Mort (Lozère). Revue d’Anthropologie, 2, 1-53.
Chikhi, L., Destro-Bisol, G.,
Bertorelle, G., Pascali, V., et Barbujani, G., 1998, Clines of nuclear DNA
markers suggest a largely Neolithic ancestry of the European gene pool.
Proceedings of the National Academy of Science USA, 95, 9053-9058.
Carlson, D. S., et Van Gerven,
D. P., 1979, Masticatory function and Post-Pleistocene in Nubia. American
Journal of Physical Anthropology, 46, 495-506.
Ciochon, R. L., Nisbett, R.
A., et Corruccini, R. S., 1997, Dietary consistency and craniofacial
development related to masticatory function in minipigs. Journal of
Craniofacial Genetics and Developmental Biology, 17, 96-102.
De Benedetto, G., Nasidze, I.
S., Stenico, M., Nigro, L., Krings, M., Vigilant, L., Stoneking, M., Paabo, S.,
et Barbujani, G., 2000, Mitochondrial DNA sequences in prehistoric human
remains from the Alps. European Journal of Human Genetics, 8, 669-677.
Ferembach, D., 1986, Les
hommes du Paléolithique supérieur autour du bassin Méditerranéen.
L´Anthropologie, 90, 579-587.
Geary, M. P. P., Pringle, J.,
Rodeck, C. H., Kingdom, J. C. P., et Hindmarsh, P. C., 2003, Sexual dimorphism
in the growth hormone and insulin-like growth factor axis at birth. The Journal
of Clinical Endocrinology and Metabolism, 88, 3708-3714.
Giesen, E. B., Ding, M.,
Dalstra, M., et van Eijden, T. M., 2003, Reduced mechanical load decreases the
density, stiffness, and strenght of cancellous bone of mandibular condyle.
Clinical Biomechanics, 18, 358-363.
Hannam, A. G., et Wood, W. W.,
1989, Relationships between size and spatial morphology of human masseter and
medial pterygoid muscles, the craniofacial skeleton, and jaw biomechanics.
American Journal of Physical Anthropology, 80, 429-445.
Irish, J. D., 2000, The
Iberomaurusian enigma: north African progenitor or dead end? Journal of Human
Evolution, 39,
393-410.
Jackes, M., Lubell, D., et
Meiklejohn, C., 1997, On physical anthropological aspects of the
Mesolithic-Neolithic transition in the Iberian Peninsula. Current Anthropology,
38, 839-846.
Kiliaridis, S., 1995,
Masticatory muscle influence on craniofacial growth. Acta Odontologica
Scandinavica, 53, 196-202.
King, R., et Underhill, P. A.,
2002, Congruent distribution of Neolithic painted pottery and ceramic figurines
with Y-chromosome lineages. Antiquity, 76, 707-714.
Lahr, M. M., 1996,
The Evolution of Modern Human Diversity. A Study of Cranial Variation.
(Cambridge: Cambridge University Press).
Lieberman,
D. E., 1996, How and why humans grow thin skulls: experimental evidence for
systemic cortical robusticity. American Journal of Physical Anthropology, 101,
217-236.
Lieberman, D. E., Pearson, O. M., et Mowbray, K. M., 2000a, Basicranial influence on overall cranial shape. Journal of Human Evolution, 38, 291-315.
Lieberman, D. E.,
Ross, C. F., et Ravosa, M. J., 2000b, The primate cranial base: ontogeny,
function, and integration. Yearbook of Physical Anthropology, 43, 117-169.
Lubell, D.,
Sheppard, P., et Gilman, A., 1992, The Maghreb. Dans Chronologies in Old World
Archaeology, vol. II, édité par R. W. Ehrich (Chicago: University Chicago
Press), pp. 257-276.
Moss, M. L., 1973, A
functional cranial analysis of primate craniofacial growth. Symposia IVth of
the International Congress of Primatology, 3, 191-208.
Moss, M. L., 1997,
The functional matrix hypothesis revisited. 4. The epigenetic antithesis and
the resolving synthesis. American Journal of Orthodontics
and Dentofacial Orthopedics, 112, 410-417.
Moss, M. L., Vilmann, H.,
Moss-Salentijn, L., Sen, K., Pucciarelli, H. M., et Skalak, R., 1987, Studies
on orthocephalization: growth behavior of the rat skull in the period 13-49
days as described by the Finite Element method. American Journal of Physical
Anthropology, 72, 323-342.
Moss, M. L., et
Young, R., 1960, A functional approach to craniology. American Journal of
Physical Anthropology, 18, 281-291.
Price, T. D., 2000, Lessons in
the transition to agriculture. Dans Europeans First Farmers, édité par D. Price
(Cambridge: Cambridge University Press), pp. 301-318.
Raadsheer, M. C., van Eijden,
T. M., van Ginkel, F. C., et Prahl-Andersen, B., 1999, Contribution of jaw
muscle size and craniofacial morphology to human bite force magnitude. Journal
of Dental Research, 78,
31-42.
Riquet, R., 1970,
Anthropologie du Néolithique et du Bronze Ancien. (Poitiers : Texier
Réunies).
Roubet, C., 2001, Neolithic of
Capsian. Dans Encyclopedia of Prehistory, vol. I Africa, édité par P. N.
Peregrine et M. Ember. (New York: Plenum Publishers), pp. 197-219.
Sardi, M. L., Novellino, P.
S., et Pucciarelli, H. M., 2003, Craniofacial morphology in the Central-West of
Argentina (Cuyo). Consequences of the transition to food production. Article
soumis.
Semino, O., Passarino, G.,
Oefner, P. J., Lin, A. A., Arbuzova, S., Beckman, L. E., De Benedictis, G.,
Francalacci, P., Kouvatsi, A., Limborska, S., Marcikiae, M., Mika, A., Mika,
B., Primorac, D., Santachiara-Benerecetti, A. S., Cavalli-Sforza, L. L., et
Underhill, P. A., 2000, The genetic legacy of Paleolithic Homo sapiens in
extant Europeans: a Y chromosome perspective. Science, 290, 1155-1159.
Sheppard, P., et Lubell, D.,
1990, Early Holocene Maghreb prehistory: and evolutionary approach. Sahara, 3, 63-69.
Simoni, L., Calafell, F.,
Pettener, D., Bertranpetit, J., et Barbujani, G., 2000, Geographic patterns of
mtDNA diversity in Europe. American Journal of Human Genetics, 66, 262-278.
Smith, P., 1979, Regional
diversity in Epipaleolithic populations. Ossa, 6, 243-247.
van Spronsen, P. H., Weijs, W.
A., Valk, J., Prahl-Andersen, B., et van Ginkel, F. C., 1991, Relationships
between jaw muscle cross-sections and craniofacial morphology in normal adults,
studied with magnetic resonance imaging. European Journal of Orthodontics, 13,
351-361.
Vogl, C., Atchley, W. C.,
Cowley, D. E., Crenshaw, P., Murray, J. D., et Pomp, D., 1993, The epigenetic
influence of growth hormone on skeletal development. Growth, Development and
Aging, 57, 163-182.
Weltman, A., Weltman, J. Y.,
Veldhuis, J. D., et Hartman, M. L., 2001, Body composition, physical exercise,
growth hormone and obesity. Eating and Weight Disorders, 6, 28-37.
Zvelebil, M., et Lillie, M.,
2000, Transition to agriculture in Eastern Europe. Dans Europeans First
Farmers, édité par D. Price, (Cambridge: Cambridge University Press), pp.
57-92.