
Olalde, I., 2023. Estudio del origen ancestral de los celtíberos con datos paleogenómicos. Antropo, 49, 1-12. www.didac.ehu.es/antropo
Artículo de revisión – Review article
Estudio del origen ancestral de los celtíberos con datos paleogenómicos
Study of the ancestral origins of the Celtiberians with palaeogenomic data
Iñigo Olalde1,2,3
1Grupo Consolidado BIOMICs, Departamento de Zoología y Biología Celular Animal, Universidad del País Vasco (UPV/EHU).
2Ikerbasque—Fundación Vasca para la Ciencia.
3Department of Human Evolutionary Biology, Harvard University; Cambridge, MA, USA.
Palabras clave: Celtíberos, Paleogenética, Next-Generation Sequencing, Cambio demográfico
Keywords: Celtiberians, Palaeogenetics, Next-Generation Sequencing, Demographic change
Resumen
A la luz de la revolución paleogenética durante la última década, la recuperación de datos genéticos nucleares de individuos antiguos está proporcionando nuevas evidencias para el conocimiento de la historia de las poblaciones humanas en el pasado. Dentro de este contexto y a pesar de la dificultad que entraña el uso generalizado de la cremación, distintos trabajos han conseguido recuperar datos genéticos de individuos procedentes de dos yacimientos de la Edad del Hierro en la periferia norte del territorio celtibérico. Junto con los 66 individuos de cronologías más antiguas secuenciados hasta la fecha en la Celtiberia, es posible llevar a cabo una primera aproximación a la ascendencia de los celtíberos. Los datos genéticos apuntan a dos cambios demográficos en esta área: el primero durante la transición Calcolítico-Edad de Bronce y el segundo al final de la Edad de Bronce, ambos involucrando un aumento de ascendencia centroeuropea. Este segundo evento, posiblemente relacionado con la llegada de hablantes de lengua celta al territorio, dio lugar a la composición genética de las poblaciones de la Edad del Hierro analizadas en este estudio, poblaciones que por otro lado mantienen claras conexiones ancestrales con las poblaciones del Bronce peninsular. En el futuro, la aplicación de las técnicas paleogenéticas a otros yacimientos del núcleo del territorio celtibérico, así como la integración de los resultados con los datos arqueológicos, lingüísticos e históricos dará como resultado un avance significativo en la comprensión de la historia de los celtíberos.
Abstract
In the light of the palaeogenetic revolution over the past decade, the retrieval of nuclear genetic data from ancient individuals is yielding new evidence for the study of human population history. Within this context and despite the difficulties imposed by the widespread use of cremation as funerary rite, several projects have generated genomic data from Iron Age individuals in the northern periphery of the Celtiberian territory. Together with the 66 individuals from older chronologies sequenced so far in this area, it is now possible to carry out an initial approximation to the ancestry of the Celtiberians. Genetic data show two demographic changes in the area: the first one during the Chalcolithic-Bronze Age transition and the second one at the end of the Bronze Age, both involving an increase of central European ancestry. The second event, possibly related to the introduction of Celtic languages in the territory, resulted in the genetic composition of the Iron Age populations analyzed in this study, populations that otherwise maintained clear ancestral connections with Bronze Age groups from the Iberian Peninsula. In the future, the application of palaeogenetic techniques to other archaeological sites located at the core of the Celtiberian territory, as well as integration of the genetic results with the available archaeological, linguistic and historical evidence will provide significant advances in the understanding of the history of Celtiberians.
Introducción
Durante la última década, la aplicación de las nuevas tecnologías de secuenciación (Next-Generation Sequencing) a la recuperación y estudio de ADN procedente de muestras antiguas está revolucionando nuestro conocimiento de la historia de las poblaciones humanas (Orlando et al, 2015). Este gran avance viene dado por la posibilidad de secuenciar cientos de miles de marcadores genéticos en el genoma nuclear, algo imposible con las técnicas previas restringidas principalmente al genoma mitocondrial, mucho más fácil de estudiar por su pequeño tamaño y alto número de copias en las células. En la actualidad, más de 10.000 genomas humanos antiguos han sido secuenciados con estas técnicas, siendo la Península Ibérica una de las regiones mejores estudiadas con cerca de 600 individuos antiguos analizados que cubren un amplio periodo desde hace 27.000 años hasta la Edad Moderna (Mallick et al., 2023).
En la Península Ibérica, uno de los períodos más interesantes para el estudio de las dinámicas poblaciones es sin duda la Edad de Hierro (desde el 900 a.C. aproximadamente hasta la conquista romana), por ser la época más antigua en la que conocemos, aunque de manera parcial e incompleta, el panorama lingüístico, es decir, la distribución de lenguas habladas en las distintas áreas y su relación filogenética. Gracias a las más de 3.000 inscripciones paleohispánicas y a la onomástica indígena en epigrafía latina (Banco de Datos Hesperia: http://hesperia.ucm.es/) podemos identificar las siguientes lenguas habladas durante la Edad de Hierro y primeros siglos de la dominación romana (Sinner y Velaza, 2019): la lengua tartésica o lengua del sudoeste, el ibérico, el lusitano, el celtibérico, el paleovasco, y una lengua indoeuropea probablemente celta en el noroeste.
De todas ellas, el celtibérico es la mejor conocida debido a tres factores: se trata de la segunda lengua paleohispánica mejor documentada después del ibérico, su sistema de escritura está razonablemente bien descifrado (Beltrán y Jordán, 2019) y al tratarse de una lengua celta de la familia indoeuropea, puede ser analizada mediante las herramientas de la lingüística comparativa. La epigrafía en lengua celtibérica se concentrada principalmente a lo largo del sistema ibérico en las provincias de Zaragoza, Teruel, Soria y Guadalajara, pero también es frecuente en las provincias limítrofes a éstas (Figura 1). No obstante, es posible que los hablantes de esta lengua se extendiesen también en regiones adyacentes pero que, al comenzar a escribir ya en lengua latina, nunca dejaran epigrafía en lengua indígena.
A pesar de que los genes no informan sobre la lengua que hablaban sus portadores, la introducción de una nueva lengua en un territorio ocurre por la llegada de hablantes de esa lengua, ya sea de forma masiva dando lugar a un cambio demográfico o a través de una élite dominante que acaba imponiendo su lengua sobre el resto. Los datos genéticos pueden, por tanto, aportar información relevante para el estudio de la historia de los hablantes de una lengua o familia de lenguas. En el caso específico de las poblaciones de habla celta durante la Edad de Hierro, existe el debate sobre el momento de su llegada en la Península, los mecanismos para su establecimiento y sus relaciones con el resto de poblaciones no celtas. La investigaciones lingüísticas, arqueológicas e históricas (Lorrio y Ruiz Zapatero, 2005) están avanzando nuestro conocimiento sobre estos aspectos, y recientemente la paleogenética promete complementar a estas disciplinas con nuevas evidencias.
Por ello, este trabajo pretende revisar los datos genéticos nucleares generados usando las técnicas de Next-Generation Sequencing a lo largo de los últimos años, con relevancia para el estudio de la historia de los celtíberos.
Materiales y métodos
Durante la última década, diversas investigaciones (Günther et al., 2015; Lipson et al., 2017; Mathieson et al., 2015; Olalde et al., 2018, 2019; Patterson et al., 2021; Valdiosera et al., 2018) han secuenciado ADN nuclear de 73 individuos antiguos procedentes de la Celtiberia, definida como el territorio con presencia significativa de epigrafía celtibérica (Figura 1). Paradójicamente, sólo siete de ellos están datados en la Edad de Hierro, mientras que el resto pertenecen a cronologías más antiguas empezando por el Neolítico Inicial ~5.100 a.C. (Anexo, Tabla A1).
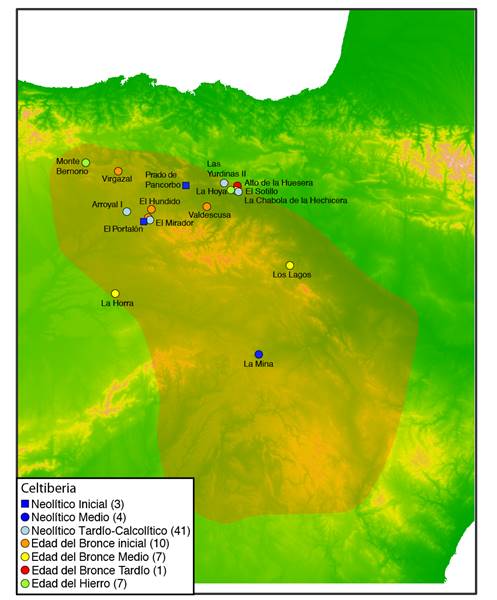
Figura 1. Mapa de la distribución geográfica de los yacimientos dentro de la Celtiberia con datos genéticos nucleares generados con técnicas de Next-Generation Sequencing. Se señala el área con presencia significativa de epigrafía celtibérica de acuerdo con el Banco de Datos Hesperia (http://hesperia.ucm.es/).
Figure 2. Map of the geographic distribution of archaeological sites within the Celtiberian territory with nuclear genetic data generated with Next-Generation Sequencing. The area with significant presence of Celtiberian epigraphy according with the Hesperia Database (http://hesperia.ucm.es/) is highlighted.
Este hecho se debe a la práctica generalizada de la cremación como ritual funerario en adultos durante la Edad de Hierro, lo que limita el número de individuos disponibles para el análisis genético a subadultos y a casos en los que no se practicó el ritual funerario habitual. De hecho, los individuos disponibles hasta el momento ilustran perfectamente ambos casos: el poblado de la Hoya que fue arrasado de manera violenta por un grupo rival y parte de sus habitantes ajusticiados (Fernández-Crespo et al, 2020; Núñez et al., 2016), y el oppidum de Monte Bernorio con presencia de neonatos enterrados bajo las casas (Torres Martínez et al., 2012). Ambos yacimientos están ubicados en el límite norte del territorio Celtibérico (Figura 1), en áreas de contacto con otros grupos étnicos como los Cántabros en el caso de Monte Bernorio y posiblemente los Vascones en el caso de La Hoya. Como consecuencia, los habitantes de estos dos yacimientos pueden no ser un muestra representativa de la ascendencia de los grupos que habitaban el núcleo del territorio celtibérico. Sin embargo, a la espera del análisis de un número más alto de individuos en zonas más centrales del territorio, estos siete individuos pueden servir como una primera aproximación a las afinidades genéticas de las poblaciones celtibéricas durante la Edad de Hierro.
Para la realización de este trabajo, se descargaron los datos genéticos nucleares de la base de datos AADR (Mallick et al., 2023) en formato Eigensoft, correspondientes a los 73 individuos antiguos disponibles en territorio celtibérico (Anexo, Tabla A1), así como los de otras poblaciones antiguas y modernas disponibles dentro y fuera de la Península.
Para analizar las afinidades genéticas de los individuos en territorio celtibérico, se realizó un Análisis de Componentes Principales con el programa smartpca del software EIGENSOFT (versión 7.2.1) (Patterson et al, 2006) sobre 591.642 SNPs autosómicos. Con el fin de evitar el efecto de la gran cantidad de marcadores sin datos en los individuos antiguos, se calcularon los componentes principales en un set 1.036 individuos modernos de Europa y Oriente Medio genotipados en el array de Human Origins (Patterson et al., 2012), y se proyectaron los individuos antiguos sobre esos componentes usando los parámetros “lsqproject:YES” and “shrinkmode:YES”.
Para modelar la ascendencia en el genoma nuclear de cada individuo, se usó el programa qpAdm del software AdmixTools v.6. (https://github.com/dReichLab/AdmixTools) (Haak et al., 2015; Harney et al, 2021). El análisis se realizó con 1.054.671 SNPs autosómicos y los siguientes outgroups: Mota, Ust_Ishim, Kostenki14, GoyetQ116-1, Vestonice16, MA1, cazadores-recolectores del este de Europa (EHG), Neolíticos de Irán, Natufienses, Yamnaya de Samara.
La determinación de los haplogrupos del cromosoma Y se llevó a cabo siguiendo la nomenclatura de ISOGG v15.73 (https://isogg.org/tree/).
Resultados
Análisis de Componentes Principales
Los resultados del Análisis de Componentes Principales para las poblaciones del área celtibérica (Figura 2) muestran una historia poblacional similar a la observada para la Península Ibérica en su conjunto (Olalde et al., 2019). Los tres individuos del Neolítico Inicial datados entre el 5.200-4.700 a.C. aparecen cerca de las poblaciones Neolíticas de Anatolia, pero desplazadas hacia las poblaciones mesolíticas de la Península. Las poblaciones del Neolítico Medio, Neolítico Tardío y Calcolítico datadas entre el 4.000-2.500 a.C. aparecen todavía más desplazadas hacia las poblaciones mesolíticas. Por último, algunos individuos pertenecientes al Calcolítico más tardío (2.500-2.200 a.C.) y las poblaciones de la Edad del Bronce y la Edad del Hierro datadas entre el 2200 a.C. y el cambio de era forman un clúster separado de las poblaciones anteriores (Figura 2). Este grupo de individuos está desplazado hacia las poblaciones de Europa Central durante la segunda mitad del tercer milenio a.C. y hacia las poblaciones de la Estepa Póntica representadas por la cultura Yamnaya hacia el 3.000 a.C. Además, es posible detectar cierta estructuración temporal dentro de este clúster, con los individuos de la Edad del Hierro algo más desplazados hacia valores más altos en el PC2. Estos resultados apuntan dos grandes cambios demográficos, uno durante la transición del Neolítico Inicial al Neolítico Medio y otro durante la transición Calcolítico-Edad de Bronce, así como a cierta discontinuidad poblacional entre la Edad de Bronce y Edad del Hierro.
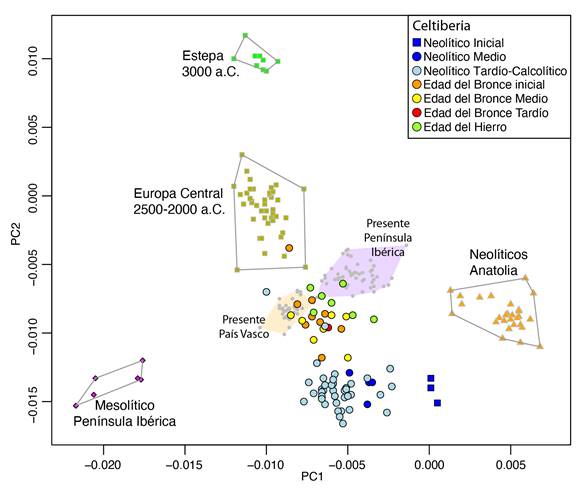
Figura 2. Análisis de Componentes Principales en 591.642 SNPs autosómicos incluyendo individuos antiguos e individuos modernos (círculos grises).
Figure 2. Principal Component Analysis on 591,642 autosomal SNPs featuring ancient individuals and present-day individuals (small grey dots).
Modelado de ascendencia en el genoma autosómico
El Análisis de Componentes Principales se trata de un análisis descriptivo de las afinidades genéticas que resulta muy útil como primera aproximación a los datos, pero no es un test formal de mezcla de poblaciones ni de discontinuidad demográfica ya que esta influido por otros procesos como la deriva genética. Por esta razón, es necesario el modelado formal de la ascendencia de los individuos, usando como fuentes de ascendencia otras poblaciones antiguas. En base a los resultados del Análisis de Componentes Principales, se propuso un modelo con tres fuentes de ascendencia: una correspondiente a cazadores-recolectores mesolíticos, otro a poblaciones Neolíticas de Anatolia y otra correspondiente a poblaciones centroeuropeas entre el 2.500-2.000 a.C. Este modelo mostró un buen ajuste a los datos genéticos en la inmensa mayoría de los individuos, y las proporciones de ascendencia derivada de cada una de las tres poblaciones de referencia puede verse en la Figura 3. Confirmando los resultados del Análisis de Componentes Principales, los individuos del Neolítico Medio, Neolítico Tardío y Calcolítico presentan ~25-30% de ascendencia de cazadores-recolectores mesolíticos, lo que implica una mayor mezcla con las poblaciones locales mesolíticas que durante el Neolítico Inicial.
A partir de la transición Calcolítico-Edad de Bronce, todos los individuos de la Celtiberia son modelados con proporciones substanciales de ascendencia centroeuropea, no presente en los periodos anteriores. Esto se explica por un fuerte impacto demográfico en la zona de las poblaciones centroeuropeas (con ancestros recientes en la Estepa Póntica) que comenzaron a llegar a la Península a partir de 2.500 a.C. De nuevo replicando lo observado en el PCA, las proporciones de ascendencia centroeuropea siguen un patrón temporal (Figura 3). Durante el Bronce Inicial la mayoría de individuos presentan un 50-70%, con excepciones notables como un individuo del yacimiento de El Hundido datado en los primeros siglos de este periodo que presenta un ~90%. Estas proporciones tienden a descender durante el Bronce Medio (1.600-1.300 a.C.) hasta valores en su mayoría por debajo del 50%, quizás debido a una mezcla con las poblaciones locales. Por último, en los individuos de la Edad de Hierro datados entre el 400 a.C. y el cambio de era se observa de nuevo un aumento de la ascendencia centroeuropea hasta el 70% en algunos casos (Figura 3).
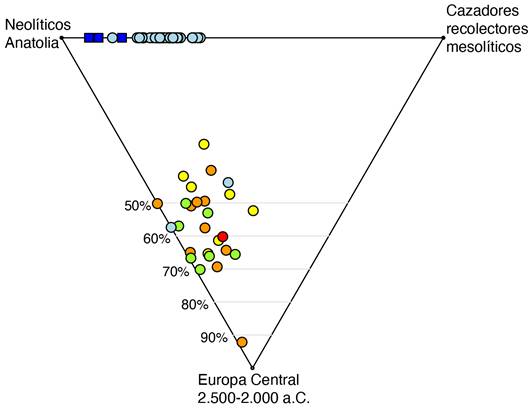
Figura 3. Modelado de la ascendencia de los individuos estudiados dentro del territorio de la Celtiberia, como una mezcla entre las poblaciones mesolíticos de cazadores-recolectores, las poblaciones neolíticas de Anatolia y las poblaciones centroeuropeas entre el 2.500-2.000 a.C. El esquema de color es el mismo que en las dos figuras anteriores.
Figure 3. Modelling the ancestry of ancient individuals from the Celtiberian territory as a mixture of Mesolithic hunter-gatherers, Neolithic Anatolians and central European populations 2,500-2,000 BCE. The color scheme is the same as in the previous two figures.
Análisis de haplogrupos del cromosoma Y
La discontinuidad demográfica observada en los datos autosómicos se ven reflejadas también en la ascendencia patrilineal determinada por el cromosoma Y. Durante los periodos Neolítico y Calcolítico se detectan hasta 6 linajes diferentes en los 24 hombres analizados, aunque con un predominio claro de I2a1b en 15 de ellos. El haplogrupo I2a1b es frecuente entre la población mesolítica europea y pudo ser incorporado a las poblaciones neolíticas a través de mezcla con las poblaciones locales. Durante la Edad de Bronce, esta diversidad desaparece y los 12 hombres pertenecen al haplogrupo R1b-M269 y, en los casos en los que la resolución lo permite, a R1b-M269-P312-DF27. Este linaje, ausente en periodos anteriores, fue introducido por los grupos centroeuropeos a principios de la Edad de Bronce, y sigue siendo el más frecuente en la actualidad en la Península Ibérica (Bejarano y Peña, 2022; Solé-Morata et al., 2017). Los linajes observados en los 3 hombres analizados en la Edad del Hierro presentan de nuevo R1b-M269 en dos de ellos (uno claramente perteneciente a DF27), pero además un caso de I2a1a1a1 (I2a-Z2049) en La Hoya. La clara presencia de I2a1a1a1 en la Península durante el Neolítico y Calcolítico (Martiniano et al., 2017; Olalde et al., 2019; Villalba-Mouco et al., 2021, 2019) hace posible que el linaje del hombre de la Hoya descienda de las poblaciones peninsulares, lo que sería un ejemplo de linaje superviviente al reemplazo por R1b-M269. Sin embargo, no es descartable que fuese introducido en la Península en algún momento a lo largo de la Edad de Bronce o Edad de Hierro desde otras áreas donde también estaba presente. En un futuro, el estudio de un número representativo de hombres durante la Edad de Hierro permitirá estimar la frecuencia este haplogrupo y determinar su origen con más precisión.
Discusión
Los resultados del Análisis de Componentes Principales y del modelado de la ascendencia indican una discontinuidad poblacional al menos parcial entre los individuos de la Edad de Hierro y las poblaciones del Bronce Medio que habitaban esos mismos territorios. Esta discontinuidad viene dada por un aumento de la ascendencia Centroeuropea en algún momento entre el ~1.300 a.C. (límite inferior de las dataciones de los individuos del Bronce Medio) y el ~380 a.C. (límite superior de la datación de los individuos de La Hoya), aunque si se tiene en cuenta el único individuo disponible del Bronce Tardío que parece presentar ya este aumento (Figura 3), se podría delimitar este evento al periodo ~1.300-900 a.C. Así pues, podemos postular tres posibles hipótesis para explicar los patrones genéticos observados en los individuos de La Hoya y Monte Bernorio:
1) En la primera, estos individuos descenderían de una mezcla entre las poblaciones locales del Bronce Medio de la zona con poblaciones centroeuropeas que llegaron al final de la Edad de Bronce.
2) En la segunda, estos individuos descenderían de una mezcla entre las poblaciones locales del Bronce Medio de la zona con poblaciones procedentes de otras áreas peninsulares con un mayor porcentaje de ascendencia centroeuropea. Las poblaciones del Bronce peninsular presentan heterogeneidad con respecto al porcentaje de ascendencia centroeuropea (Villalba-Mouco et al., 2021), por lo que no es necesario invocar la llegada directa de poblaciones centroeuropeas.
3) En la tercera, estos individuos descenderían directamente de poblaciones procedentes de otras áreas peninsulares con un mayor porcentaje de ascendencia centroeuropea, sin contribución de las poblaciones locales anteriores.
En cualquier caso, la contribución de poblaciones peninsulares, ya sea de la misma Celtiberia o de otras regiones parece necesaria a la vista de los datos genómicos autosómicos y de la presencia en Monte Bernorio del linaje R1b-DF27, característico de las poblaciones peninsulares durante la Edad del Bronce.
Finalmente, estos resultados nos permiten identificar dos posibles momentos para la llegada de los hablantes de la lengua celtibérica a la Celtiberia, suponiendo que dicha llegada vino acompañada de un cambio demográfico suficientemente grande como para dejar huella en el registro paleogenético. El primer momento sería durante la transición Calcolítico-Edad de Bronce ~2.200 a.C., acompañando el gran cambio demográfico observado en la Península y en toda Europea occidental (Haak et al., 2015; Olalde et al., 2018) que supuso la llegada de poblaciones esteparias y su mezcla con las poblaciones locales. Este evento es muy probablemente el responsable de la introducción inicial de las lenguas indoeuropeas en Europa (Willerslev et al, 2023), lo que en principio cuadraría con la posible introducción del celtibérico (una lengua indoeuropea de la rama celta) y otras lenguas hispano-célticas en la península. No obstante, este modelo entraría en conflicto con las dataciones lingüísticas del proto-céltico más tardías, que lo sitúan durante la transición Bronce-Hierro. El segundo posible momento sería al final de la Edad de Bronce, acompañando al aumento de ascendencia centroeuropea que con los datos disponibles podemos delimitar ~1.300-900 a.C, y coincidiendo con un importante cambio demográfico recientemente documentado en el sur de Gran Bretaña (Patterson et al., 2021) que ha sido postulado como un posible vector para la expansión de las lenguas celtas en ese territorio.
En conclusión, el análisis del genoma nuclear de los individuos de la Edad del Hierro procedentes de La Hoya y Monte Bernorio nos brinda una primera aproximación a la ascendencia de los celtíberos, con las debidas reservas que derivan del bajo tamaño muestral y de la localización periférica de ambos asentamientos con respecto al núcleo del territorio de la Celtiberia, y a la espera de futuros estudios que con toda probabilidad proporcionarán evidencias más robustas.
Referencias
Bejarano, Y. y Peña, J. A. 2022. Origen de la distribución geográfica del haplogrupo R-M269. Antropo, 48, 41–52.
Beltrán, F. y Jordán, C. 2019. Writing and language in Celtiberia. In Palaeohispanic Languages and Epigraphies. Oxford University Press. https://doi.org/10.1093/oso/9780198790822.003.0010
Fernández-Crespo, T., Ordoño, J., Llanos, A., et al. (2020). Make a desert and call it peace: massacre at the Iberian Iron Age village of La Hoya. Antiquity, 94(377), 1245–1262. https://doi.org/10.15184/aqy.2020.161
Günther, T., Valdiosera, C., Malmström, H., et al. 2015. Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques. Proceedings of the National Academy of Sciences of the United States of America, 112, 11917–11922. https://doi.org/10.1073/pnas.1509851112
Haak, W., Lazaridis, I., Patterson, N., et al. 2015. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature, 522, 207–211. https://doi.org/10.1038/nature14317
Harney, É., Patterson, N., Reich, D., et al. 2021. Assessing the performance of qpAdm: A statistical tool for studying population admixture. Genetics, 217(4). https://doi.org/10.1093/GENETICS/IYAA045
Lipson, M., Szécsényi-Nagy, A., Mallick, S., et al. 2017. Parallel ancient genomic transects reveal complex population history of early European farmers. Nature, 551, 368–372. https://doi.org/10.1101/114488
Lorrio, A. J., y Ruiz Zapatero, G. 2005. The Celts in Iberia: an overview. E-Keltoi Journal of Interdisciplinary Celtic Studies, 6, 167–254. Retrieved from http://rua.ua.es/handle/10045/20403
Mallick, S., Micco, A., Mah, M., et al. 2023. The Allen Ancient DNA Resource (AADR): A curated compendium of ancient human genomes. BioRxiv : The Preprint Server for Biology, (Figure 1), 0–7. https://doi.org/10.1101/2023.04.06.535797
Martiniano, R., Cassidy, L. M., Ó’Maoldúin, R., et al. 2017. The population genomics of archaeological transition in west Iberia: Investigation of ancient substructure using imputation and haplotype-based methods. PLoS Genetics, 13(7), e1006852. https://doi.org/10.1371/journal.pgen.1006852
Mathieson, I., Lazaridis, I., Rohland, N., et al. 2015. Genome-wide patterns of selection in 230 ancient Eurasians. Nature, 528, 499–503. https://doi.org/10.1038/nature16152
Núñez, C., Baeta, M., Cardoso, S., et al. 2016. Mitochondrial DNA Reveals the Trace of the Ancient Settlers of a Violently Devastated Late Bronze and Iron Ages Village. Plos One, 11(5), e0155342. https://doi.org/10.1371/journal.pone.0155342
Olalde, I., Brace, S., Allentoft, M. E., et al. 2018. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature, 555(7695), 190–196. https://doi.org/10.1038/nature25738
Olalde, I., Mallick, S., Patterson, N., et al. 2019. The genomic history of the Iberian Peninsula over the past 8000 years. Science, 363(6432), 1230–1234. https://doi.org/10.1126/science.aav4040
Orlando, L., Gilbert, M. T. P., y Willerslev, E. 2015. Reconstructing ancient genomes and epigenomes. Nature Reviews Genetics, 16(7), 395–408. https://doi.org/10.1038/nrg3935
Patterson, N., Isakov, M., Booth, T., et al. 2021. Large-scale migration into Britain during the Middle to Late Bronze Age. Nature, 601(7894), 588–594. https://doi.org/10.1038/s41586-021-04287-4
Patterson, N., Moorjani, P., Luo, Y., et al. 2012. Ancient admixture in human history. Genetics, 192(3), 1065–1093. https://doi.org/10.1534/genetics.112.145037
Patterson, N., Price, A. L., y Reich, D. 2006. Population structure and eigenanalysis. PLoS Genetics, 2(12), e190. https://doi.org/10.1371/journal.pgen.0020190
Sinner, A. G., y Velaza, J. 2019. Palaeohispanic Languages and Epigraphies. Oxford University Press. https://doi.org/10.1093/oso/9780198790822.001.0001
Solé-Morata, N., Villaescusa, P., García-Fernández, C., et al. 2017. Analysis of the R1b-DF27 haplogroup shows that a large fraction of Iberian Y-chromosome lineages originated recently in situ. Scientific Reports, 7(1), 1–13. https://doi.org/10.1038/s41598-017-07710-x
Torres Martínez, J. F., Martínez Velasco, A., De Luis Mariño, S., et al. 2012. El oppidum de Monte Bernorio en la Cantabria Histórica. Nueve siglos de Historia. Kobie Serie PaleoantroPología, 31, 137–156.
Valdiosera, C., Günther, T., Vera-Rodríguez, J. C., et al. 2018. Four millennia of Iberian biomolecular prehistory illustrate the impact of prehistoric migrations at the far end of Eurasia. Proceedings of the National Academy of Sciences of the United States of America, 115(13), 201717762. https://doi.org/10.1073/pnas.1717762115
Villalba-Mouco, V., Oliart, C., Rihuete-Herrada, C., et al. 2021. Genomic transformation and social organization during the Copper Age-Bronze Age transition in southern Iberia. Science Advances, 7(47), 22–27. https://doi.org/10.1126/sciadv.abi7038
Villalba-Mouco, V., van de Loosdrecht, M. S., Posth, C., et al. 2019. Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula. Current Biology, 29, 1–9. https://doi.org/10.1016/j.cub.2019.02.006
Willerslev, E., Kroonen, G., y Kristiansen, K. (Eds.). 2023. The Indo-European Puzzle Revisited: Integrating Archaeology, Genetics and Linguistics. Cambridge: Cambridge University Press.
Anexo
Tabla A1. Tabla resumen de los datos genéticos de los 73 individuos antiguos estudiados en territorio celtibérico.
Table A1. Summary table of the genetic data from 73 ancient individuals from Celtiberian territory.
|
ID |
Cronología |
Yacimiento |
SNPs autosómicos recuperados |
Sexo genético |
Haplogrupo cromosoma Y (notación ISOGG v15.73) |
Haplogrupo mitocondrial |
|
I2199 |
NeIn |
El Prado de Pancorbo |
322693 |
F |
H1 |
|
|
I1972 |
NeIn |
El Prado de Pancorbo |
361966 |
F |
K1a4a1 |
|
|
atp005 |
NeIn |
El Portalón |
75023 |
M |
G2a2a |
J2b1a |
|
I0405 |
NeMe |
La Mina |
329685 |
M |
H2c1a |
K1a1b1 |
|
I0406 |
NeMe |
La Mina |
759352 |
M |
I2a1b1a2b |
H1 |
|
I0407 |
NeMe |
La Mina |
562183 |
F |
K1b1a1 |
|
|
I0408 |
NeMe |
La Mina |
906735 |
F |
U5b1 |
|
|
I3276 |
NeTa-Ca |
Alto de la Huesera |
711562 |
M |
G2a2a1 |
V |
|
I3277 |
NeTa-Ca |
Alto de la Huesera |
114489 |
F |
U5b2b |
|
|
I1843 |
NeTa-Ca |
Alto de la Huesera |
642124 |
F |
U5b1 |
|
|
I1981 |
NeTa-Ca |
Alto de la Huesera |
116962 |
F |
J1c1 |
|
|
I1845 |
NeTa-Ca |
Alto de la Huesera |
403825 |
M |
I2a1b1b1a1 |
K1a2c |
|
I1846 |
NeTa-Ca |
Alto de la Huesera |
649096 |
F |
T2b3+151 |
|
|
I1978 |
NeTa-Ca |
Alto de la Huesera |
846240 |
F |
K1a4a1 |
|
|
I2473 |
NeTa-Ca |
El Sotillo |
151260 |
M |
I2a1b |
H3 |
|
I1976 |
NeTa-Ca |
El Sotillo |
135886 |
M |
I2a1b1 |
H3 |
|
I2467 |
NeTa-Ca |
El Sotillo |
123929 |
M |
I2a1b1 |
X2 |
|
I3273 |
NeTa-Ca |
La Chabola de la Hechicera |
129303 |
F |
J2a1a1 |
|
|
I3272 |
NeTa-Ca |
La Chabola de la Hechicera |
177439 |
M |
I2a1b1b |
X2b+226 |
|
I1975 |
NeTa-Ca |
La Chabola de la Hechicera |
103350 |
M |
I2a1b |
J1c |
|
I1838 |
NeTa-Ca |
Las Yurdinas II |
828092 |
F |
J1c3 |
|
|
I1842 |
NeTa-Ca |
Las Yurdinas II |
450494 |
M |
G2a2a |
H1e1a |
|
I3269 |
NeTa-Ca |
Las Yurdinas II |
656625 |
M |
I2a1b1b |
U5b2b3a |
|
I3270 |
NeTa-Ca |
Las Yurdinas II |
276586 |
F |
J2a1a1 |
|
|
I3271 |
NeTa-Ca |
Las Yurdinas II |
320045 |
F |
K1a4a1 |
|
|
I1281 |
NeTa-Ca |
El Mirador |
623387 |
F |
H1t |
|
|
I1272 |
NeTa-Ca |
El Mirador |
578482 |
F |
K1b1a |
|
|
I1271 |
NeTa-Ca |
El Mirador |
231181 |
F |
K1a+195 |
|
|
I1276 |
NeTa-Ca |
El Mirador |
113000 |
F |
H3c3 |
|
|
I1280 |
NeTa-Ca |
El Mirador |
201692 |
F |
J1c1 |
|
|
I1282 |
NeTa-Ca |
El Mirador |
67061 |
M |
I2 |
H3 |
|
I1284 |
NeTa-Ca |
El Mirador |
121241 |
M |
I2a1b |
H3 |
|
I1300 |
NeTa-Ca |
El Mirador |
778557 |
F |
K1a2a |
|
|
I1303 |
NeTa-Ca |
El Mirador |
449619 |
M |
I2a1a1a1a1 |
U3a1 |
|
I1314 |
NeTa-Ca |
El Mirador |
215998 |
M |
G2a2b2b1a1a |
J2a1a1 |
|
I5838 |
NeTa-Ca |
El Mirador |
698649 |
M |
I2a1b1b1a1 |
K1a+195 |
|
I1277 |
NeTa-Ca |
El Mirador |
543744 |
M |
I2a1b1b |
H3 |
|
por002 |
NeTa-Ca |
El Portalón |
500379 |
M |
I2a1a2 |
K1a2b |
|
atp016 |
NeTa-Ca |
El Portalón |
1148173 |
F |
X2c2 |
|
|
por004 |
NeTa-Ca |
El Portalón |
82801 |
M |
I2a1b1 |
K1a |
|
atp12-1420 |
NeTa-Ca |
El Portalón |
946761 |
M |
I2a1b1b |
H3c |
|
ATP17 |
NeTa-Ca |
El Portalón |
22396 |
M |
I2a1b |
H3 |
|
atp002 |
NeTa-Ca |
El Portalón |
1146940 |
M |
H2c1a~ |
U5b3 |
|
ATP20 |
NeTa-Ca |
El Portalón |
13065 |
F |
U5a1c |
|
|
I0459 |
NeTa-Ca |
Arroyal I |
521175 |
F |
U5b2b |
|
|
I0460 |
NeTa-Ca |
Arroyal I |
330551 |
M |
I2a1b1b |
H45 |
|
I0461 |
NeTa-Ca |
Arroyal I |
715168 |
F |
K1a1b1 |
|
|
I0462 |
NeTa-Ca |
Arroyal I |
32133 |
F |
K1a+195 |
|
|
EHU002 |
BrIn |
El Hundido |
662713 |
M |
R1b1a1b1a1a2 |
K1a4a1 |
|
EHU001 |
BrIn |
El Hundido |
535632 |
M |
R1b1a1b |
U5a1b1 |
|
I5665 |
BrIn |
Virgazal |
476556 |
M |
R1b1a1b1a1a2 |
K1a24a |
|
I6470 |
BrIn |
Virgazal |
674668 |
M |
R1b1a1b1a1a2a |
J1c1 |
|
VAD002 |
BrIn |
Valdescusa |
94921 |
M |
R1b1a1 |
J2b1a2 |
|
VAD003 |
BrIn |
Valdescusa |
33906 |
F |
HV0 |
|
|
VAD004 |
BrIn |
Valdescusa |
276491 |
M |
R1b1a1b1a1a |
J1c3 |
|
VAD001 |
BrIn |
Valdescusa |
560204 |
M |
R1b1a1b1a1a2a5 |
U5b1 |
|
VAD005 |
BrIn |
Valdescusa |
386287 |
M |
R1b1a1b1a1 |
J2b1a2 |
|
ATP9 |
BrIn |
El Portalón |
279955 |
F |
U5b1b |
|
|
I2471 |
BrMe |
El Sotillo |
197375 |
F |
K1b1a |
|
|
I1840 |
BrMe |
El Sotillo |
649557 |
M |
R1b1a1b1a1a |
H3ap |
|
I1977 |
BrMe |
El Sotillo |
116516 |
F |
U5b1 |
|
|
I2472 |
BrMe |
El Sotillo |
143324 |
M |
R1b1a1b1a1 |
K1a+195 |
|
I2470 |
BrMe |
El Sotillo |
866444 |
M |
R1b1a1b1a1a2a |
J2a1a1 |
|
I3490 |
BrMe |
El Cerro |
211552 |
F |
HV0b |
|
|
esp005 |
BrMe |
Cueva de los Lagos |
903470 |
M |
R1b1a1b1a1a2a |
K1a |
|
I2469 |
BrTa |
El Sotillo |
214996 |
F |
H3an |
|
|
I3757 |
Hi |
La Hoya |
121747 |
F |
J1c1 |
|
|
I3758 |
Hi |
La Hoya |
859014 |
F |
U5b1 |
|
|
I3759 |
Hi |
La Hoya |
723750 |
M |
I2a1a1a1 |
H1 |
|
I19989 |
Hi |
Monte Bernorio |
49891 |
F |
H1e1a6 |
|
|
I19991 |
Hi |
Monte Bernorio |
383018 |
M |
R1b1a1b1a1a2a |
U5b1g |
|
I19990 |
Hi |
Monte Bernorio |
110087 |
M |
R1b1a1b1a1a2 |
U5b1g |
|
I19987 |
Hi |
Monte Bernorio |
309026 |
F |
H1e1a6 |
NeIn: Neolítico Inicial; NeMe: Neolítico Medio; NeTa-Ca: Neolítico Tardío-Calcolítico; BrIn: Edad del Bronce Inicial; BrMe: Edad del Bronce Medio; BrTa: Edad del Bronce Tardío; Hi: Edad del Hierro.