
Girotti, M., De Iasio, S., Orrù, A., Boano, R.,
Boetsch, G. 2017. Opportunità di selezione naturale in una comunità alpina
dell’Antico Delfinato. Antropo, 38, 1-12. www.didac.ehu.es/antropo
Opportunità di selezione naturale in una
comunità alpina dell’Antico Delfinato
Opportunity for Natural Selection in an
Alpine community of Antico Delfinato
M.
Girotti1, S. De Iasio2, A. Orrù1, R. Boano1,
G. Boetsch3
1
Dip. Scienze della Vita e Biologia dei Sistemi. Università di Torino (Italia)
2
Dip. Scienze Chimiche, della Vita e
della Sostenibilità Ambientale. Università di Parma (Italia)
3
UMIESS3189 (CNRS/UCAD/UGB/USTTB/CNRST). Dakar.
Parole
chiave: selezione naturale, Indice di Crow,
popolazioni alpine
Key
words: natural selection, Crow's Index, alpine
population
Abstract
Nelle popolazioni umane il successo riproduttivo dipende dal
numero totale di discendenti che ogni coppia o meglio, ogni donna, lascia alle
generazioni successive. Tuttavia la selezione naturale non opera solo
attraverso la fecondità differenziale, in quanto la fitness biologica è
determinata anche dalla sopravvivenza dei discendenti, almeno fino al
raggiungimento della loro età riproduttiva.
Il presente studio è stato condotto sulla popolazione di
Chiomonte, piccolo comune rurale della Val di Susa (Piemonte, Italia). Sono
state analizzate le variazioni dell’indice di opportunità di selezione naturale
di Crow (It) e delle sue componenti (indici di mortalità preriproduttiva
Im e di fecodità If) dal 1670 al 1830. In
questo intervallo di tempo nella comunità si sono verificati importanti cambiamenti
che, a livello economico e sociale, hanno avuto ripercussioni su alcuni aspetti
relativi alla struttura delle famiglie e alla loro discendenza.
A partire dagli atti di
matrimonio, battesimo e decesso riportati nei registri parrocchiali è stato
possibile ricostituire le famiglie “biologiche” e analizzare la storia
riproduttiva di circa l’80% delle donne sposate, valutando il numero di figli
avuti da ciascuna in relazione all’età al matrimonio e alla sua durata. L’analisi
dell’interazione tra i patterns di mortalità e l’opportunità di selezione
naturale ha permesso di comprendere le potenziali implicazioni delle crisi di
mortalità e soprattutto il ruolo che la mortalità infantile aveva in passato. Questa,
infatti, si mantiene su valori prossimi al 250‰ fino alla seconda metà del
XVIII secolo per poi abbassarsi a valori mediamente inferiori al 220‰, comportando
una diminuzione del peso della componente di mortalità preriproduttiva sulla
opportunità di selezione totale. Nel trend di variazione di It, infatti,
si possono identificare due periodi: un primo, dal 1670 fino alla metà del
XVIII secolo, nel quale It presenta un andamento relativamente
costante e un secondo in cui tende a diminuire progressivamente. Negli ultimi
periodi, infine, si osserva la tendenza a un progressivo aumento della
componente legata alla fecondità.
Abstract
The
reproductive success in human populations depends on the total number of
descendants that each couple (or each woman) leaves to subsequent generations.
Natural
selection does not work only through differential fertility, in fact biological
fitness is also determined by the survival of the descendants, at least until
their reproductive age is reached.
The
present study was conducted on the population of Chiomonte, a small rural
village in the Susa Valley (Piedmont, Italy). We analyzed variations of the Crow’s index
of opportunity for natural selection (It)
and its components (Im and
If) from 1670 to 1830. During this timeframe, major economic and social changes occurred in the
community, which have had repercussions on certain aspects of the structure of
the families and on their descendants.
Drawing
from marriage, baptism, and death acts recorded in the parish registers, it was
possible to reconstruct "biological" families in order to analyse the
reproductive history of about 80% of the married women, by assessing the number
of children each of them had in relation to their age at marriage and its
duration.
The
analysis of the interaction between mortality models and the opportunity for
natural selection has allowed us to understand the potential implications of
mortality crises and, above all, the role that infant mortality had in the past. This remains at values up to 250 ‰ until the second half of the
eighteenth century and then reduces to mean values below 220 ‰, with a
consequent reduction of the importance of the pre-reproductive mortality
component on total selection.
Two
periods can be identified in the trend of It's
variation: first, from 1670 until the middle of the eighteenth century, where
it remains relatively constant, and a second in which it tends to decrease progressively.
Furthermore, we observed a trend of progressive increase in the fertility
component.
Introduzione
La selezione naturale, che è alla base della teoria evoluzionistica
darwiniana, è uno dei più importanti fattori che provocano cambiamenti nel pool
genico delle popolazioni umane. Essa esplica la sua funzione mediante l’azione
di due meccanismi demografici: la sopravvivenza differenziale e la fecondità
differenziale. La sopravvivenza differenziale garantisce agli individui più
idonei di rimanere in vita almeno fino all’età riproduttiva, mentre gli
“inidonei” muoiono prima; tale meccanismo può essere visto anche nell’ottica
opposta, cioè di “mortalità differenziale”. La fecondità differenziale consente
ai più idonei di lasciare una prole più numerosa e ai meno idonei di lasciare
meno figli o non lasciarne affatto: il successo riproduttivo di differenti
genotipi dipende dal numero totale di figli per coppia - o, meglio, per donna -
lasciati alle generazioni successive (Jacquard, 1970).
L’analisi dei trend nella
struttura demografica di una popolazione è un valido strumento di indagine
micro-evolutiva e le variazioni nel tempo di parametri demografici, come la
fecondità e la mortalità preriproduttiva, possono riflettere processi di
selezione naturale ed essere di grande interesse negli studi di genetica di
popolazione (Terrenato et al., 1979;
Alfonso-Sanchez et al., 2004).
Nelle moderne società occidentali, caratterizzate da alti livelli di
industrializzazione e urbanizzazione, grandi miglioramenti nelle condizioni
socio-economico-culturali, igienico sanitarie e sviluppo della moderna medicina
preventiva hanno determinato una drastica riduzione della mortalità preriproduttiva
e in particolare di quella infantile e neonatale (Zonta et al., 1997; Ulizzi et al.,
1998; Astolfi et al., 2000). Rispetto
al periodo preindustriale infatti le risposte alle fluttuazioni ambientali
risultano più efficaci; la pressione della selezione dovrebbe quindi essersi
ridotta con il trascorrere del tempo, fenomeno conosciuto come “relaxation of natural selection” (Ulizzi
and Terrenato 1987, 1992).
Le società preindustriali, invece, sono state caratterizzate
da ricorrenti crisi demografiche, risultato di un ambiente più instabile e
imprevedibile (Palloni et al., 1988).
È generalmente riconosciuto che epidemie, carestie e guerre rappresentino i più
importanti fattori nel determinare riduzione e cambiamento strutturale di una
popolazione (Del Panta, 1980; Alfonso-Sánchez et al., 2004). Durante questi
episodi catastrofici, cambiamenti delle caratteristiche demografiche (mortalità
preriproduttiva, fecondità, nuzialità e movimenti migratori, che spesso si
discostano largamente dai loro livelli di equilibrio) possono riflettersi sui
processi selettivi (Terrenato et al.,
1979; Reddy and Chopra, 1990; Ulizzi et
al., 1998). Queste variabili operano singolarmente o in sinergia per
determinare la fitness di una particolare popolazione in un ambiente specifico (Mohan
Rao et al., 2015). Inoltre esse hanno
una forte influenza sulle conseguenze genetiche a lungo termine (Bittles, 1994;
Mielke and Pitkänen, 1989).
Le crisi di popolazioni sono spesso avvenute in passato quando
la disponibilità dei dati demografici era frammentaria o mancante; pertanto le
informazioni riguardanti la mortalità per sesso o età (che sono di grande
utilità per valutare le conseguenze biologiche di una crisi) sono spesso incomplete
oppure inesistenti (Watkins and Menken, 1985; Palloni et al., 1988; Mielke and Pitkänen, 1989). Le implicazioni
demografiche, genetiche ed evolutive, correlate alle crisi preindustriali, non sono
quindi ancora adeguatamente comprese soprattutto a causa della scarsità di
statistiche ragionevolmente affidabili. È estremamente importante, per quanto
possibile, esaminare l’insieme dei dati, l'intensità e l'impatto delle crisi
nel contesto storico del momento; inoltre dovrebbe essere data particolare
attenzione a quelle situazioni critiche per le quali esiste una documentazione
ampia e precisa (Pitkänen et al.
1989; Del Panta and Rettaroli, 1994; Alfonso-Sanchez et al., 2004).
La severità di una crisi
dipende da due fattori, entrambi correlati alla struttura della popolazione per
sesso ed età: la sua resilienza e la dimensione delle perdite subite. Una crisi
che colpisce le classi più giovani e quelle in età riproduttiva avrà
conseguenze che sono più gravi nel medio e nel lungo termine - per le stesse
perdite complessive – di una che interessa soprattutto le persone anziane
(Del Panta, 1980). Studiarne gli effetti sulle dinamiche demografiche significa
anche valutare il peso delle diverse componenti della selezione naturale; sebbene
sia difficile misurarne l'intensità direttamente nelle popolazioni umane, è possibile
valutarne gli effetti, anche da un punto di vista puramente demografico, analizzando
le tendenze riproduttive in una generazione di donne oppure attraverso il
calcolo dell’indice di opportunità
della selezione naturale (Crow, 1958). Tale indice, sebbene non possa
essere considerato di tipo analitico in quanto non dà informazioni sulla
relazione tra un particolare genotipo e la sua fitness, permette di valutarne i
cambiamenti mediante specifici tassi di natalità e mortalità (Alfonso Sanchez et al., 2004). Questo metodo di calcolo,
definito come una funzione di due componenti demografiche, fornisce il limite
superiore del valore dell’opportunità selettiva che esiste in ogni popolazione
e potrebbe misurare l’effettiva selezione solo se le ereditabilità delle
fitness fossero complete e se i tassi di natalità e mortalità dipendessero
totalmente da fattori genetici. Nonostante tali limiti, l’indice di Crow è
stato largamente usato negli studi di popolazioni umane. Il metodo fornisce una
media di confronto tra la fecondità differenziale e la mortalità, fenomeni
necessari - ma non sufficienti - affinché avvenga la selezione. Poterlo calcolare, in totale assenza
dell’ammontare della popolazione e della sua struttura per età, è
fondamentale perché è molto difficile disporre dell’una o dell’altra:
occorrerebbe essere in possesso di almeno uno stato delle anime o un censimento,
documentazione non comune per le popolazioni del XVII-XVIII secolo (Redy and Chopra,
1990; Lakshmi et al., 2005; Prakash
and Narayanan, 2009).
Materiali e
metodi
Questo studio è stato
condotto sulla popolazione di Chiomonte, un piccolo comune rurale della Val di
Susa (Piemonte, Italia) situato a circa 750 m slm. Nel periodo analizzato
(1670-1829), il numero di residenti non superava i 1500 abitanti. Grazie alla
sua strategica posizione geografica, al confine tra Delfinato e Ducato di
Savoia, per secoli le attività commerciali furono molto sviluppate (Casalis,
1837). Dal punto di vista storico Chiomonte appartenne all’antico Delfinato
fino al 1713 quando, al termine della guerra di successione spagnola, passò
sotto il Ducato di Savoia. L’influenza d’Oltralpe continuò comunque a farsi sentire
considerevolmente almeno fino all’Unità d’Italia.
Chiomonte dispone di una
ricca documentazione relativa agli atti di battesimo, di matrimonio e di
sepolture, risalente alla seconda metà del XVII secolo. È stato quindi
possibile effettuare la ricostruzione delle famiglie biologiche a partire dagli
atti costitutivi di matrimonio, associando a ciascuno di essi le storie
individuali dei figli (Fleury and Henry, 1976). Le dinamiche riproduttive di
ciascuna donna sono state raccolte nelle fiches
des familles che hanno costituito la base per lo studio della mortalità e
della fecondità necessarie per il calcolo dell’indice di Crow.
Per il periodo 1670-1829
sono stati analizzati 9223 atti di battesimo, 8941 di sepoltura e 2035 di
matrimonio. Le famiglie ricostruite sono state riunite in otto coorti
ventennali aventi come origine la data di celebrazione del primo matrimonio.
Per l'analisi della
mortalità è stata considerata, oltre alla mortalità in generale, anche quella
infantile (precedente il compimento del primo anno di vita) e quella giovanile o
preriproduttiva (entro il 16° anno di età).
La fitness è stata valutata
considerando il numero complessivo di figli avuti da ciascuna donna; sono stati
selezionati sia i matrimoni delle spose alle prime nozze sia le eventuali
unioni successive e sono state considerate solo le spose per le quali è stato
reperito l’atto di morte, così da avere notizie certe sulla durata del
matrimonio e sull’esplicarsi del potenziale riproduttivo.
È stato calcolato il numero medio di figli delle donne che
hanno completato il ciclo riproduttivo (età alla morte > 45 anni) mettendolo
in relazione alla durata del matrimonio e all’età della donna alla prima unione.
Sono stati esclusi i
pochi casi di donne per le quali non è stato possibile attribuire il numero di
figli in quanto sono stati ritrovati solo gli atti di decesso dei bambini e non
quelli di nascita; si tratta delle cosiddette naissances perdues, cioè bambini nati o battezzati in altre
parrocchie e che sfuggono al conteggio (Schiaffino, 1978).
Opportunità della selezione naturale
Per valutare l’opportunità
della selezione naturale è stato utilizzato l’indice di Crow (1958):
It = Im + (1/Ps)If
dove:
Im = Pd/ Ps (Indice di mortalità
preriproduttiva), essendo
Pd = proporzione di individui deceduti in età preriproduttiva
(< 16 anni)
Ps = proporzione di individui che hanno raggiunto l’età adulta
(≥ 16 anni)
If = δ2 /μ2 (Indice di
fecondità) dove:
δ2 = varianza del numero dei nati vivi del periodo
μ = numero medio dei nati vivi del periodo
Poiché non è stato
possibile reperire l’atto di decesso nel 10% circa dei figli inseriti nelle famiglie
ricostruite It è stato calcolato in due modi: nel primo considerando
solo i sopravvissuti certi ai 16 anni, per i quali è stato rinvenuto l’atto di
decesso; nel secondo, considerando sopravvissuti tutti i giovani per i quali
non è stato trovato l’atto di decesso, ipotizzando che avessero raggiunto l’età
adulta e fossero emigrati.
Risultati
Popolazione
Dall’analisi degli atti
di nascite, decessi e matrimoni è stato possibile riconoscere importanti
differenze nel numero degli eventi tra un anno e l’altro (Fig. 1).
Per quanto riguarda i
decessi è possibile riconosce le crisi di mortalità che hanno colpito la
popolazione; la più importante di queste è conseguente a “febbri maligne” che si manifestarono in Alta
Val di Susa dall’autunno del 1690 alla fine del 1692 in occasione del passaggio
di truppe francesi e che decimarono molte persone (Baccon Bouvet, 1999). A Chiomonte l’epidemia colpì inizialmente i soldati a partire dalla fine
di ottobre 1690 e da dicembre si diffuse a tutta la popolazione prolungandosi
sino al maggio del 1691. Si
tratta di una grave crisi in quanto
l’incremento percentuale di decessi supera il 300% della media degli eventi dei
periodi non interessati da crisi (Del Panta, 1980).
Le nascite, pur
presentando anch’esse notevoli variazioni puntuali, tendono ad aumentare
progressivamente; alla fine del XVIII secolo si osserva un importante
incremento e nel primo ventennio del XIX secolo risultano sempre mediamente
superiori ai decessi.
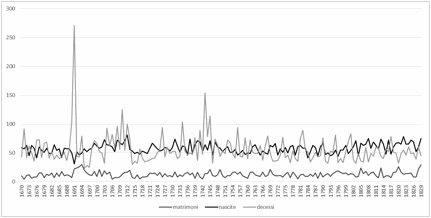
Figura 1. Matrimoni, nascite e
decessi.
Figure 1. Marriages, births and
deaths.
Le vicende storiche e le
economie locali hanno avuto importanti ripercussioni sul comportamento matrimoniale.
Chiomonte, per la sua posizione geografica e per gli scambi commerciali,
risulta aperta fin dal XVII secolo, infatti mediamente oltre il 20% delle
unioni sono esogamiche.
Si osservano importanti
decrementi delle unioni nei periodi di crisi di mortalità dovuti a epidemie o a
momenti di difficoltà conseguenti a eventi bellici. L’aumento delle unioni, una
volta passato il periodo critico, rappresenta la classica risposta delle
popolazioni a una crisi di mortalità: particolarmente importante è stato, ad
esempio, il picco osservato dopo la grave epidemia del 1690-1690. Nonostante
questi comportamenti e il conseguente aumento delle nascite il saldo naturale
si mantiene negativo fino alla fine del XVIII secolo anche se negli ultimi
decenni si manifestano i segnali di un cambiamento di tendenza.
Mortalità infantile e giovanile
Come si può osservare
nella figura 2 i morti infanti rappresentano in media un quarto dei decessi
totali per tutto il periodo considerato, ad eccezione del ventennio 1690-1709, quando
scendono a circa il 20%, a seguito di una importante crisi di mortalità avvenuta
tra la fine del 1690 e l’inizio del 1691 che ha maggiormente inciso sulla
frazione adulta della popolazione (Boëtsch et
al., 2007). Leggermente più elevata è la mortalità dei giovani tra 1 e 15
anni con percentuali che superano anche il 30% dei decessi totali. Complessivamente
la mortalità infanto-giovanile costituisce oltre il 50% dei decessi.
La forte diminuzione
percentuale dei morti infanti nel periodo 1690-1709 è conseguente alla grave
crisi del 1690-1691. Durante
questo periodo il livello generale della mortalità aumenta e la percentuale di
bambini al di sotto dell’anno si riduce proporzionalmente al 10%, mentre
risultano molto superiori le altre classi di età. È una crisi di mortalità che
colpisce particolarmente la componente giovanile e dei giovani adulti, con un
forte impatto negativo sulla velocità di ripresa della popolazione.
La mortalità infantile
presenta una lenta ma progressiva diminuzione seppur con qualche fluttuazione
sebbene, fino al 1770, si mantenga su valori prossimi al 250‰. Dalla seconda
metà del XVIII secolo si abbassa ulteriormente presentando valori mediamente
inferiori al 220‰.
Come si può notare dalla
figura 3 la mortalità neonatale (decessi avvenuti entro il primo mese di vita),
anche se con differenze più o meno marcate, è sempre superiore alla post
neonatale (decessi tra il secondo e il dodicesimo mese di vita).
Relativamente alla
mortalità neonatale risulta che la componente precoce rappresenta sempre oltre
il 50% degli eventi.
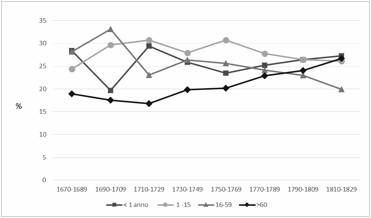
Figura 2. Mortalità per fasce di
età.
Figure 2. Mortality by age
groups.
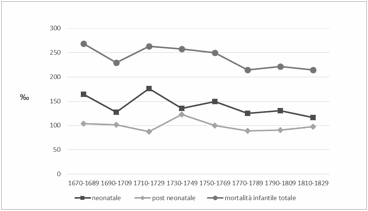
Figura 3. Mortalità infantile e
sue componenti.
Figure 3. Infant mortality and its components.
Ricostruzione famiglie e fitness
In regime di fecondità
naturale il numero dei figli è legato all’età della madre al matrimonio e alla
durata del matrimonio stesso. A Chiomonte la maggior parte delle donne si sposava
tra i 20 e 24 anni. Il numero medio dei figli passa da circa 2 per le unioni
che durano meno di 10 anni a 6 per le unioni che durano più di 20 anni (Tab.1).
|
Anni |
0-9 anni |
0-9 anni* |
10-19 anni |
10-19 anni* |
>20 anni |
>20 anni* |
|
1670-1689 |
1,90 |
2,30 |
4,50 |
4,96 |
6,36 |
6,74 |
|
1690-1709 |
1,66 |
2,29 |
4,18 |
4,77 |
6,70 |
7,21 |
|
1710-1729 |
2,54 |
3,14 |
4,81 |
5,38 |
6,38 |
7,19 |
|
1730-1749 |
1,97 |
2,48 |
4,43 |
4,68 |
6,00 |
6,34 |
|
1750-1769 |
1,79 |
2,26 |
4,76 |
5,03 |
6,33 |
6,58 |
|
1770-1789 |
2,00 |
2,52 |
3,97 |
4,56 |
5,98 |
6,44 |
|
1790-1809 |
1,74 |
2,04 |
4,16 |
5,16 |
5,72 |
5,98 |
|
1810-1829 |
2,00 |
2,12 |
3,86 |
4,77 |
5,76 |
6,13 |
Tabella 1. Numero medio di figli in
relazione alla durata del matrimonio.
*numero medio calcolato
escludendo le donne che non hanno avuto figli
Table 1. Average number of children related to the marriage length.
* average number
calculated excluding women who did not have children
Il numero medio di figli
per donna (escludendo quelle che dovrebbero non averne avuti) aumenta
progressivamente nei primi tre ventenni (Fig.4), passando da 4,7 a 6,2; successivamente
cala, fluttuando tra 5,3 e 5,8 a testa.
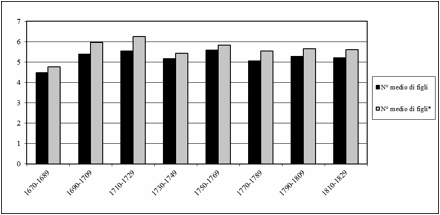
Figura 4. Numero medio di figli
per donna.
* numero medio calcolato escludendo le donne che
non hanno avuto figli.
Figure 4. Average number of children per woman.
* average number
calculated excluding women who did not have children.
La fitness presenta un progressivo
aumento. Le donne che hanno almeno un figlio che raggiunge l’età riproduttiva,
sono poco più del 70% nel 1670-1689; aumentano progressivamente, dall’83% all’88%
nei cento anni successivi fino a raggiungere oltre il 90% dal 1790 (Tab. 2).
|
Anni |
Donne con figli |
Donne con discendenza |
Donne senza discendenza |
Donne con discendenza ignota |
|
1670-1689 |
153 |
110 (71,90%) |
29 (18,95%) |
14 (9,15%) |
|
1690-1709 |
178 |
148 (83,15%) |
18 (10,11%) |
12 (6,74%) |
|
1710-1729 |
138 |
116 (84,06%) |
14 (10,14%) |
8 (5,80%) |
|
1730-1749 |
160 |
135 (84,37%) |
16 (10,00%) |
9 (5,63%) |
|
1750-1769 |
160 |
136 (85,00%) |
11 (6,88%) |
13 (8,12%) |
|
1770-1789 |
146 |
128 (87,78%) |
9 (6,16%) |
9 (6,16%) |
|
1790-1809 |
173 |
157 (90,75%) |
13 (7,51%) |
3 (1,73%) |
|
1810-1829 |
191 |
174 (91,10%) |
8 (4,19%) |
9 (4,71%) |
Tabella
2 . Fitness delle
donne.
Table
2 . Women's
Fitness.
Opportunità di selezione
Le tabelle 3 e 4 mostrano
i valori dell’indice di Crow (It) e delle componenti Im,
If e If/Ps.
Il trend di variazione
di It sembra identificare due periodi: un primo periodo dal 1670
fino alla metà del XVIII secolo, nel quale It presenta un andamento
relativamente costante e un secondo periodo caratterizzato da una progressiva
diminuzione; il valore si riduce di oltre il 50% dal 1670-1689 al 1810-1829 se
consideriamo solo i sopravvissuti certi (Tab. 3), mentre nel secondo caso (Tab.
4) It risulta decisamente inferiore e la differenza tra il primo e
l’ultimo periodo è meno marcata.
Una migliore
comprensione del trend di It si ottiene analizzandone le componenti.
Considerando le tabelle
3 e 4 si osserva che la frazione Pd si mantiene su valori prossimi
allo 0,5 fino alla metà del 1700, per poi diminuire progressivamente a partire
dal terzo periodo considerato con un decremento totale del 30%.
Mentre il numero medio di figli µ per
donna con il tempo tende ad assumere un valore costante, la differenza tra la
varianza e tale valore è sempre alta, ad eccezione del periodo 1790-1809 dove è
di 1,08, e raggiunge il valore massimo di 4,1 nel 1730-1749 (Tab. 3-4).
I valori di Im
scendono da 1,628 del 1670-1689 a 0,767 nell’ultimo periodo. Se consideriamo
come sopravvissuti tutti i bambini di cui non si è trovato il decesso Im
passa da 1,233 a 0,628. Im presenta un trend del tutto conforme a
quello di It come evidenziato dalla correlazione altamente
significativa (Fig. 5 - 6).
|
Indice di Crow |
1670-1689 |
1690-1709 |
1710-1729 |
1730-1749 |
1750-1769 |
1770-1789 |
1790-1809 |
1810-1829 |
|
|
Mortalità |
ps |
0,34 |
0,36 |
0,35 |
0,35 |
0,42 |
0,41 |
0,51 |
0,51 |
|
pd |
0,55 |
0,51 |
0,55 |
0,54 |
0,48 |
0,48 |
0,41 |
0,39 |
|
|
Im |
1,63 |
1,41 |
1,58 |
1,55 |
1,16 |
1,17 |
0,79 |
0,77 |
|
|
Fecondità |
N |
153,00 |
178,00 |
138,00 |
160,00 |
160,00 |
146,00 |
173,00 |
191,00 |
|
μ |
4,76 |
5,96 |
6,25 |
5,43 |
5,84 |
5,53 |
5,64 |
5,61 |
|
|
σ2 |
7,44 |
9,30 |
9,34 |
9,53 |
8,04 |
7,96 |
6,72 |
7,65 |
|
|
If |
0,33 |
0,26 |
0,24 |
0,32 |
0,24 |
0,26 |
0,21 |
0,24 |
|
|
Totale |
If/ps |
0,97 |
0,72 |
0,68 |
0,93 |
0,57 |
0,63 |
0,41 |
0,48 |
|
It |
2,60 |
2,13 |
2,26 |
2,47 |
1,73 |
1,79 |
1,20 |
1,25 |
Tabella 3. Indice di Crow calcolato considerando
solo i sopravvissuti certi.
Table 3. Crow
index calculated considering only the certain survivors.
|
Indice di Crow |
|
1670-1689* |
1690-1709* |
1710-1729* |
1730-1749* |
1750-1769* |
1780-1789* |
1800-1809* |
1810-1829* |
|
Mortalità |
ps |
0,45 |
0,49 |
0,45 |
0,46 |
0,52 |
0,52 |
0,59 |
0,62 |
|
|
pd |
0,55 |
0,51 |
0,55 |
0,54 |
0,48 |
0,48 |
0,41 |
0,39 |
|
|
Im |
1,23 |
1,05 |
1,23 |
1,17 |
0,94 |
0,93 |
0,68 |
0,63 |
|
Fecondità |
N |
153,00 |
178,00 |
138,00 |
160,00 |
160,00 |
146,00 |
173,00 |
191,00 |
|
|
μ |
4,76 |
5,96 |
6,25 |
5,43 |
5,84 |
5,53 |
5,64 |
5,61 |
|
|
σ2 |
7,44 |
9,30 |
9,34 |
9,53 |
8,04 |
7,96 |
6,72 |
7,65 |
|
|
If |
0,33 |
0,26 |
0,24 |
0,32 |
0,24 |
0,26 |
0,21 |
0,24 |
|
Totale |
If/ps |
0,73 |
0,54 |
0,53 |
0,70 |
0,46 |
0,50 |
0,36 |
0,39 |
|
|
It |
1,97 |
1,59 |
1,76 |
1,87 |
1,39 |
1,43 |
1,04 |
1,02 |
Tabella 4. Indice di Crow calcolato considerando
come sopravvissuti anche coloro per il quali non si è trovato l’atto del
decesso.
Table 4. Crow
index calculated considering as survivors even those for which we did not find
the act of death.
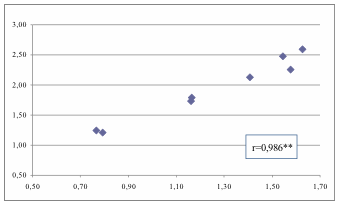
Figura 5. Im vs It
calcolato considerando solo i giovani che sicuramente hanno raggiunto i 16 anni
di età.
Figure 5. Im vs It calculated considering only the
youngsters who certainly reached the age of 16.
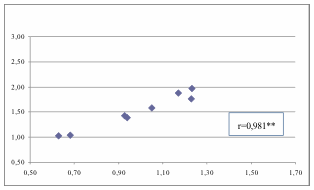
Figura
6. Im vs It calcolato considerando tutti i giovani che
hanno raggiunto i 16 anni di età, compresi quelli di cui non è stato trovato
l’atto di decesso.
Figure
6.
Im vs It
calculated considering all youngsters who reached the age of 16, including those
for which we did not find the act of death.
Relativamente costante
risulta invece la componente If che, salvo i due ventenni iniziali
in cui è poco superiore a 0,3, mentre negli altri oscilla tra 0,21 e 0,26.
Nella tabella 5 sono riportati i contributi relativi di Im e di If
rispetto a It (%Im/It
e % If/It).
Il contributo
corrispondente della mortalità differenziale sulla selezione è considerevole in
tutto il periodo ed è sempre superiore al 60%.
Il contributo relativo di If è
scarso: poco oltre il 12% tra il 1670 e il 1709, scende al 10,5% nel 1710-1729,
dopo di che aumenta progressivamente per raggiungere il valore massimo di 19,5%
nell’ultimo ventennio.
|
Anni |
Im |
If |
If* |
|
1670-1689 |
62,71 |
12,65 |
16,70 |
|
1690-1709 |
66,15 |
12,33 |
16,51 |
|
1710-1729 |
69,77 |
10,58 |
13,55 |
|
1730-1749 |
62,48 |
13,10 |
17,29 |
|
1750-1769 |
67,24 |
13,65 |
16,91 |
|
1770-1789 |
65,06 |
14,44 |
18,12 |
|
1790-1809 |
65,77 |
17,53 |
20,34 |
|
1810-1829 |
61,45 |
19,45 |
23,75 |
Tabella
5. Contributi relativi di Im e If
su It (%Im/It
e % If/It).
Table
5. Im and If related
contributions on It
Come si può osservare
dalla figura 7, i valori di It sono in relazione inversa rispetto
alla frazione Ps.
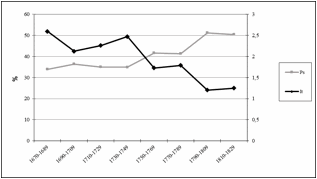
Figura
7. Relazione tra It e Ps
Figure
7. Relationship
between It and Ps
L’esclusione dei giovani
per i quali non è stato reperito l’atto di decesso porta senz’altro ad una
sovrastima dei valori di It, mentre considerarli come sopravvissuti
oltre i 16 anni ne comporta una sottostima, poiché non si può escludere che una
certa percentuale di questi non sia sopravvissuta: il più attendibile valore di
It è quindi un valore intermedio tra i due, più spostato verso l’uno
o verso l‘altro, in relazione alla reale sopravvivenza all’età riproduttiva.
Discussione
La struttura dei
matrimoni, l'età dei coniugi, la mortalità e la fecondità differenziale, la
consanguineità e le migrazioni sono i fattori fondamentali nell’evoluzione di
una popolazione.
Nel periodo considerato
Chiomonte si presenta con le caratteristiche classiche di una comunità di
Ancien Régime di piccole dimensioni: il numero dei matrimoni - e
conseguentemente delle nascite - varia considerevolmente da un anno all’altro,
anche in relazione a eventi contingenti.
A Chiomonte i cambiamenti
che, a livello economico e sociale, si sono verificati tra il 1670 e il 1830, hanno
avuto ripercussioni su alcuni aspetti relativi alla struttura e alla
discendenza delle famiglie. L'aumento della durata media della vita ha
comportato anche un prolungamento della durata dei matrimoni che ha determinato
a sua volta, come prima conseguenza, un incremento del numero di donne che alla
fine del matrimonio avevano raggiunto e superato i 45 anni, portando a termine
il loro periodo riproduttivo.
Come riscontrato anche in altre comunità
(Navarra, 1998) tanto più è giovane la donna al momento del matrimonio tanto
maggiore è la possibilità di avere un numero più elevato di figli. Nei periodi
studiati, infatti, non sono validi i modelli riproduttivi che caratterizzano le
popolazioni attuali.
Il ruolo dei fattori più
legati ad aspetti genetici e meno influenzati da quelli ambientali può essere
dedotto valutando il peso della mortalità neonatale, che statisticamente è
dovuta prevalentemente a fattori endogeni. Essa
rappresenta sempre oltre il 26% degli eventi, con un massimo di oltre 30% nel 1799-1809.
Pur supponendo che non tutti i decessi avvenuti in questa fascia di età siano
dovuti esclusivamente a fattori endogeni, indubbiamente quelli genetici
rivestono un ruolo importante sull’ammontare della mortalità preriproduttiva.
I valori di It
e delle sue componenti riscontrati nel nostro campione sono paragonabili a
quelli osservati in Piemonte nel 1871 e nel periodo 1872-81 (Terrenato et al., 1979). I valori osservati a
Chiomonte anticipano di circa 50 anni quelli globali del Piemonte: questo dato
è compatibile con una più bassa mortalità infantile osservata in ambiente di
montagna rispetto ai centri di pianura (Viazzo, 1994). Alti valori di It
sono noti in popolazioni storiche caratterizzate da alta mortalità infantile;
quando questa comincia a diminuire e aumenta la componente fecondità, It
tende ad abbassarsi. In Italia l’indice si è ridotto del 75% nel corso del XIX secolo;
il suo valore, osservato a Chiomonte, nell’ultimo periodo risulta inferiore a
quello (1,960) riferito all’Italia nel 1871 (Terrenato et al., 1979). Anche in zone rurali dell’Europa centrale tra il
1765 e il 1899 sono stati osservati valori di It paragonabili a quelli
del nostro campione (Nowak et al.,
2013).
I valori di If
risultano invece molto bassi rispetto a quelli osservati in Piemonte nel 1871
(0,651) (Terrenato et al., 1979). Nel
nostro caso If presenta poche
variazioni, nonostante il numero medio di figli presenti delle oscillazioni, in
quanto la varianza si mantiene sempre alta. If diventa determinante solo quando il livello di
mortalità infantile è molto basso, quindi quando il valore di Im scende.
L’importanza di If
è osservata in altre comunità: quella basca, dove tale indice presenta un alto
contributo anche se solo a partire dall’inizio del XX secolo (Alfonso-Sanchez et al., 2004); in comunità attuali
dell’Andhra Pradesh (Lakhmi et al.,
2005; Hed, 1987); in due gruppi di russi, dove la componente che caratterizza
la fecondità differenziale (If) supera quella della mortalità
infantile (Im) (Tabikhanova and Osipova, 2012); in altre comunità
indiane dove migliori condizioni di vita e di salute sono state indicate come
possibile causa di un valore di If considerevolmente maggiore di
quello di Im (Sohklhlet, 2013).
It si abbassa
progressivamente in quanto il peso della componente rappresentata dalla
frazione di mortalità preriproduttiva tende a diminuire. Negli ultimi periodi
la componente legata alla fecondità comincia ad aumentare: si hanno le prime
avvisaglie dei cambiamenti che si verificano nelle società, legati al
miglioramento delle condizioni che porteranno questo fattore ad assumere sempre
maggiore rilevanza, fino a diventare attualmente l'elemento determinante del
valore dell'indice di opportunità di selezione naturale.
Sulla base dei risultati ottenuti possiamo concludere che la popolazione in
esame è stata caratterizzata da una alta selezione naturale, influenzata sia
dai fattori culturali che da quelli biologici, che ne ha fortemente
condizionato lo sviluppo.
Fino fine del XVIII secolo le nascite infatti non riescono a
compensare le gravi perdite subite in conseguenza delle crisi di mortalità; il saldo
naturale risulta ancora negativo sebbene nell’ultimo ventennio del XVIII secolo
si verifichi una inversione di tendenza: sono le prime avvisaglie della
transizione demografica (Chesnais, 1986). Il numero di figli per donna si
mantiene relativamente costante e alto per tutto il periodo analizzato, siamo
infatti in un’epoca caratterizzata da alta fecondità conseguente allo scarso o
nullo controllo delle nascite. Solo quando comincia a calare la mortalità
infantile e un numero più alto di soggetti raggiunge l’età riproduttiva, con
conseguente diminuzione di It, la popolazione tende ad aumentare: il
fattore positivo legato alla natalità riesce a compensare quello negativo
rappresentato dalla mortalità.
La diminuzione della mortalità preriproduttiva e in
particolare di quella infantile, seppur debole, rappresenta una delle
conseguenze dell’inizio del miglioramento delle condizioni ecologiche,
economiche e sociali che caratterizza i paesi europei all’inizio del XIX secolo.
Bibliografia
Alfonso-Sánchez M.A., Calderon R., Peña J.A., 2004, Opportunity for natural selection in a Basque
population and its secular trend: evolutionary implications of epidemic
mortality. Human Biology, 76, 3, 361-381.
Astolfi P., Ulizzi L., Zonta L.A., 2000, Natural
selection and reproductive behavior in Italy 1930-1993. Human Biology, 72,
349-357.
Baccon Bouvet C.,
1999, Salbertrand – Storia di una comunità alpina e della sua Valle. (Melli, Borgone di Susa -TO).
Bittles A. H., 1994, The role and significance of consanguinity as a
demographic variable. Population and development review, 20,
3, 561-184.
Boëtsch G., Girotti M., Rabino Massa E., Piguel A., 2007, Comparaison entre
le profil démographique d’une épidémie de grippe à Chiomonte en 1690-1691
(Turin Italie) avec d’autres crises démographiques (peste, choléra). Peste: entre
épidémies et sociétés. (Firenze University press), 149-153.
Casalis G.,
1837, Dizionario geografico degli Stati Sardi. Vol IV (Maspero, Mazzorati e
comp.).
Chesnais J.C., 1986, La transition démographique. Etapes,
formes, implications économiques. Travaux et documents, Cahier 113.
Crow F.J., 1958, Some possibilities for measuring
selection intensities in man. Human Biology, 30,1-13.
Del
Panta L., 1980, Le epidemie nella storia demografica italiana (secoli XIV-XIX).
(Loescher Editore, Torino).
Del Panta L.,
Rettaroli R., 1994, Introduzione alla demografia storica. (Laterza,
Bari).
Fleury M., Henry L., 1976, Nouveau manuel de
dépouillement et d’exploitation de l’état civil ancien, INED, Paris.
Hed H.M.E., 1987, Trends in Opportunity for natural
selection in the Swedish population during the period 1650-1980. Human Biology, 59,5, 785-797.
Jacquard A,. 1970, Structures génétiques des populations.
(Masson INED, Paris).
Lakhmi N., Venkateswara RaoT., Veerraju P., 2005,
Opportunity for natural selection among three endogamous subpopulations of
Andhra Pradesh. Indian Journal of Human Genetics, January-April, 11, 1, 39-43.
Mielke J. H., Pitkänen K. J., 1989, War demography: The impact of the
1808-09 war on the civilian population of Åland, Finland. Eur. J. Popul. 5, 373-398.
Mohan Rao P., Ramesh M., Sudhakar G., 2015, Measuring
opportunity for natural selection in two endogamous sub populations of Andhra
Pradesh, South India. International Journal of Current Research, 7, 05, 15554-15558.
Navarra
E., 1998, Demografia di un villaggio alpino della Carnia: nuzialità e natalità
a Sauris tra Settecento e Ottocento. La ricerca
folklorica, 38, 49-61.
Nowak O., Liczbińska G., Piontek J., 2013,Opportunity
for the Operation of Natural Selection in a Contemporary Local Population (The
Case of Slovincians, Poland), Advances in Anthropology, 3, 3, 121-126.
Palloni A., Komlos J., Watkins S. C., Menken J., 1988,
On the role of crises in historical perspective: An exchange. Popul. Dev. Rev.,
14, 145-170.
Pitkänen K.J., Mielke J. H,. Jorde L.B., 1989,
Smallpox and its eradication in Finland: Implications for disease control. Popul. Stud., 42, 95-111.
Prakash D.S.R.S., Narayanan S., 2009, Opportunity for
natural selection in Yerukula tribe of coastal Andhra Pradesh. Stud Tribes
Tribals, 7, 1, 67-69.
Reddy B.M., Chopra V.P., 1990, Occupation and the
opportunity for natural selection: the Indian case. International Journal of
Anthropology. 3, 4, 295-308.
Schiaffino A.
1978. Le « naissances perdues » nella ricostruzione di famiglie su
base parrocchiale. Statistica XXXVIII, 2: 251-262
Sohkhlet B., 2013, Index of opportunity for natural
selection among the Gowdas of Kodagahalli village, Karnataka, India Indian J
Hum Genet. Jul-Sep; 19(3), 315–319.
Tabikhanova L. E., Osipova L. P., 2012, Analysis of the genetic and demographic structure
of populations from Aginskii Buryat district contrasting in habitation
conditions, Russian Journal of Genetics, 48, 12, 1239–1246.
Terrenato L., Ulizzi L., San Martini A., 1979, The
effects of demographic transition on the opportunity for selection: changes
during the last century in Italy. Ann. Hum. Gen., 42, 391-399.
Ulizzi L., Astolfi P., Zonta L.A., 1998, Natural
selection in industrialized countries: A study of three generations of Italian
newborns. Ann. Hum Genet., 62, 47-53.
Ulizzi L., Terrenato L., 1987, Natural selection
associated with birth weight. V. The secular relaxation of the stabilizing
component. Ann. Hum Genet., 51, 205-210.
Ulizzi L., Terrenato L., 1992, Natural selection
associated with birth weight. VI. Towards the end of the stabilizing component.
Ann. Hum Genet., 56, 370-376.
Viazzo P.P., 1994, Les modèles alpins de la mortalité
infantile. Annales de Démographie Historique, 97-117.
Watkins S.C., Menken J., 1985, Famines in historical perspective.
Population and Development Review. 11, 4, 647-75.
Zonta L.A., Astolfi P., Ulizzi L., 1997, Heterogeneous
effects of natural selection on the Italian newborns. Ann. Hum Genet., 61,
137-142.