
Cocilovo, J.A., Varela, H.H., Fuchs, M.L., 2017. Divergencia fenotípica y
flujo génico en el Área Andina Centro Sur: el trayecto entre Puna de Jujuy, San
Pedro de Atacama y Calama. Antropo, 37, 1-15. www.didac.ehu.es/antropo
Divergencia fenotípica y flujo génico en el Área
Andina Centro Sur: el trayecto entre Puna de Jujuy, San Pedro de Atacama y
Calama
Phenotypic divergence and gene flow in the Andean Central South Area:
the route between Puna de Jujuy, San Pedro de Atacama and Calama
José A. Cocilovo,
Héctor H. Varela y María L. Fuchs
Departamento de Ciencias Naturales, Facultad
de Ciencias Exactas, Físico-Química y Naturales, Universidad Nacional de Río
Cuarto, 5800 Río Cuarto, Argentina.
Correspondencia: José A. Cocilovo Departamento de Ciencias
Naturales, Facultad de Ciencias Exactas, Físico-Química y Naturales,
Universidad Nacional de Río Cuarto, 5800 Río Cuarto, Argentina. E-mail: jcocilovo@exa.unrc.edu.ar
Palabras claves: poblaciones prehistóricas, deriva genética, flujo genético, caracteres
cuantitativos, FST
Key words: prehistoric population, genetic drift, gene flow, quantitative traits, FST.
Resumen
Un tema de gran interés en la
investigación antropológica es el estudio de las relaciones entre las poblaciones
del Área Andina Centro Sur basadas en el impacto producido por el intercambio de
conocimientos, de ideas y de productos materiales en la subsistencia y la
consolidación de grupos locales. Dicho proceso debió haber involucrado también
el apareamiento entre individuos de distintas localidades y regiones
(migraciones), circunstancia que influyó en la composición genética y en la
diferenciación de las poblaciones. En el presente trabajo se evalúa la magnitud
de dicha interacción y su influencia en la divergencia genética ancestral entre Calama (CAL, 1390 d.C.), San Pedro de Atacama
(SPA, 400 a.C. y 1450 d.C.) y la Puna de
Jujuy (PJ, 1029 y 1497 d.C.). En 1107 individuos se midieron 31 variables craneométricas.
Para establecer la estructura de la población y evaluar las relaciones
genéticas entre los grupos se aplicaron métodos estadísticos de análisis multivariado
y el coeficiente Fst para la heredabilidad de 0,55. Los resultados permiten
inferir diferencias fenotípicas estadísticamente significativas entre los grupos (Wilks'
Lambda: 0,26315 aproximación F(62,2148) = 32,892, p<0,01). El estadístico Fst = 0,063 ± 0,002074 indica la magnitud de la divergencia y el nivel
del flujo génico entre subáreas. La variación genética existente es el producto
de un antiguo proceso evolutivo que dominó una vasta región del Área Andina
Centro-Sur configurando el perfil genético de las poblaciones a nivel local y
regional.
Abstract
A subject of great interest in anthropological research is the study of the relations between the populations of the Andean Area South Center based on the impact produced by the exchange of knowledge and material products in the subsistence and consolidation of local groups. This process should have also involved mating between individuals from different localities and regions (migrations), that influenced the genetic composition and the differentiation of populations. In the present work the magnitude of this interaction and its influence on the genetic ancestral divergence is evaluated between Calama (CAL, 1390 AD), San Pedro de Atacama (SPA, 400 BC y 1450 AD) and Puna de Jujuy (PJ, 1029 y 1497 AD). In a sample of 1107 individuals, 31 craniometric variables were measured. To establish the population structure and to evaluate the genetic relationships between the groups, statistical methods of multivariate analysis and the Fst coefficient for the heritability of 0.55 were applied. The results allow inferring statistically significant phenotypic differences between the groups (Wilks' Lambda: 0.26315 F(62,2148) approximation = 32.892, p <0.01). The statistic Fst = 0.063 ± 0.002074 show the magnitude of the divergence and the level of the gene flow between subareas. The existing genetic variation is the product of an old evolutionary process that dominated the Andean Central-South Area, configuring the genetic profile of populations at local and regional level.
Introducción
En el Área Andina Centro Sur la exploración, conquista y ocupación de
nuevos espacios aptos para el desarrollo de la vida fue el resultado de un
extenso proceso adaptativo durante el cual factores genéticos y culturales
determinaron la diversidad fenotípica que caracteriza a distintas regiones y
localidades. El poblamiento inicial fue protagonizado por grupos pequeños que
se dispersaron por un amplio y accidentado territorio ocupando econichos aptos
para la vida. La supervivencia estuvo basada en la eficiencia para la captación
de recursos y en la existencia de vastas redes relacionadas con el intercambio de
productos y de personas. La distribución de sitios arcaicos en el Área Andina
Centro Sur (Aldenderfer y Flores Blanco, 2011) es el resultado de este proceso
dispersivo. Por milenios y más de 400 generaciones, durante la conquista y
colonización de nuevos territorios aptos para la subsistencia, los grupos
humanos experimentaron cambios en su composición genética por variaciones en el
tamaño efectivo y en el éxito reproductivo.
La fluctuación de la señal antropogénica basada en el registro de marcas
radiocarbónicas refleja los avatares de la población regional. En efecto, el
estudio de este registro permitió identificar fases en las cuales se observan
caídas de la señal (7.500-7.900 cal a.P. y entre 6.900-6.300 cal a.P.) y otros
un incremento de la señal (11.100 a.P. cal y a.P. de 4.700 cal). Tales eventos podrían
indicar fluctuaciones en la densidad de la población, la cual habría
experimentado un incremento importante entre el Holoceno Medio y Tardío (Muscio
y López, 2016). Es posible que dichas fluctuaciones determinaran cambios en la
composición genética de las poblaciones locales y su diferenciación.
Por ejemplo, el efecto principal de la disminución de la población es la
trasmisión no aleatoria de los genes de una generación a la siguiente
determinando la pérdida de variabilidad y el incremento de la homocigosis por endogamia
y deriva génica. Además existen otros dos fenómenos asociados: el efecto
“cuello de botella”, cuando una población experimenta una reducción del tamaño
efectivo entre generaciones y el “efecto fundador” cuando un grupo pequeño se
desprende de una población mayor y abandona una localidad para colonizar otro
espacio apto para la subsistencia. En estos casos se produce un incremento de
la homocigosidad y la divergencia genética de la población ancestral (Hartl y
Clark, 1997). En general la variabilidad genética necesaria para la
subsistencia es mantenida por el equilibrio entre la deriva y el aporte de
novedades genéticas por migración. El éxito en la captación y manejo de
recursos naturales junto a una mayor interacción regional permitió el
incremento de la población y el restablecimiento de la diversidad genética (heterocigosidad).
La distancia es un factor importante por el cual se produce la extinción
del parentesco y la diferenciación fenotípica entre localidades de distintas regiones
(Morton 1982; Barbujani 1987). Este factor fue explorado a partir del estudio
de muestras antiguas de Perú, Chile y Argentina (Cocilovo, 1981; Cocilovo y Di
Rienzo, 1984-1985; Cocilovo y Rothhammer, 1990; Cocilovo y Varela, 1999). Posteriormente,
se obtuvo una explicación más precisa del proceso evolutivo al medir el efecto combinado de la deriva genética y de
la migración en la diferenciación de las poblaciones. Esta información además permitió
establecer las relaciones de ancestralidad más probables a nivel local y
regional (Varela et al, 2008, 2013 y
2014) incluyendo el sur de Bolivia, Chile y Argentina.
La investigación arqueológica revela la ocupación del territorio a
partir de 13.000-12.000 años a.C. y el
desarrollo de distintas etapas, períodos o fases que explican la evolución de
la sociedad y la interacción entre distintas subáreas. Dicha interacción
incluyó el tráfico de productos y el tránsito de personas a través de una
compleja red de caminos y de rutas que conectaron localidades y regiones de
Perú, Bolivia, Norte de Chile y Noroeste Argentino hasta tiempos históricos
(Tarragó, 1977; Núñez y Dillehay, 1995; Nielsen, 2002, 2011; Llagostera, 1995; Montenegro y
Ruiz, 2007; De Nigris, 2009; Yacobaccio,
2012, Molina Otárola, 2011, etc).
San Pedro de Atacama (SPA) es una localidad del Norte Árido de Chile
ubicada a 2.436 msnm, con un clima árido, escasas precipitaciones, intensa
radiación solar y gran amplitud térmica. En esta localidad, se registran tumbas
con contenidos funerarios del acervo
material e iconográfico local y foráneo (Tarragó 1977, 1984, 1989; Llagostera
1995, Llagostera et al. 1988). La
presencia de individuos migrantes fue detectada en base a las marcas
establecidas por la concentración de 87Sr/86Sr en los sitios de Coyo-3, Coyo
Oriental, Solcor-3 y Larache (Knudson et
al. 2012, Torres Rouff et al. 2015).
Un caso testigo sobre la presencia foránea fue determinado en una tumba del
ayllu de Coyo Oriental conteniendo cuatro mujeres con evidencias de lesiones
provocadas por leishmaniasis, una infección adquirida en zonas trasandinas
(Costa et al. 2009, Costa y
Llagostera 2014).
El estudio de la variabilidad genética mitocondrial de PJ revela la
existencia haplogrupos únicos y compartidos entre distintos sitios del Área
Andina Centro Sur. En PJ se determinó la frecuencia dominante del linaje A2 con
una gran diversidad haplotípica. Además se registraron otros linajes
compartidos con poblaciones actuales y antiguas de Costa de Perú, Quebrada de
Humahuaca, Tucumán, Río Negro, Mbyá-Guaraní, Tupí, etc. (Postillone et al. 2017). Igualmente en la Quebrada
de Humahuaca se comprobó también una alta frecuencia del haplogrupo A2 y cinco
haplotipos idénticos o semejantes a los haplotipos fundadores distribuidos por
todas las regiones de Sudamérica, incluyendo las llanuras orientales del Chaco,
Argentina y Brasil. El análisis realizado sobre haplogrupos del cromosoma Y
reveló su pertenencia a poblaciones Amerindias, entre los cuales uno de los haplotipos
fue observado en la vertiente Andina
oriental de Bolivia (provincia de Beni), otro en Toba (Chaco,
Argentina), otro en Quichuas de Ecuador, cuatro en Uros de Perú y nueve
haplotipos fueron idénticos a los encontrados en una muestra de Perú y mientras
que la menor distancia estimada fue con comunidades Guaraní, Salta, Criollos y
Chaco de Argentina (Mendisco et al.
2014). Además la frecuencia de 25% de A2 en una muestra de Atacameños y de B2 de
70 % en Aymara y 60 % en Atacameño (San Pedro de Atacama, Norte de Chile) y el
valor Fst: 0,070 (p = 0,027) entre ambas etnias (de Saint Pierre et al. 2012) permite inferir la
señal de una antigua relación entre NOA,
Bolivia y SPA coincidiendo con la investigación arqueológica. Además, recientes
investigaciones permitieron comprobar en particular las relaciones entre PJ y
SPA (Fuchs et al. 2016b, Cocilovo et al. 2017).
Considerando la información proporcionada por la investigación
arqueológica, etnohistórica, antropológica y genética molecular, el objetivo
del presente trabajo es analizar la estructura de la población y el efecto
migratorio en la diferenciación entre Calama (CAL), San Pedro de Atacama (SPA)
y la Puna de Jujuy (PJ) a partir de rasgos fenotípicos. Tales localidades
constituyen, tres espacios claves del desarrollo humano antiguo en el Área
Andina Centro Meridional por las relaciones entre el Norte de Chile, el
Noroeste Argentino, sur de Perú, sur de Bolivia y bosques tropicales.
Materiales y Métodos
La muestra de SPA proviene de varios sitios estudiados anteriormente
(Varela et al. 1990; Cocilovo et al. 1994; Cocilovo et al. 1995; Varela y Cocilovo 1996;
Varela y Cocilovo 2009; etc). Los materiales empleados corresponden al
intervalo temporal entre el 300 a.C. y 1.532 d.C. incluyendo los períodos que
se mencionan a continuación: el período Intermedio Temprano o Formativo está
integrado por las fases Toconao y Sequitor (300 a.C. - 400 d.C.), entre el 300 d.C.
y el 1.000 d.C. se desarrolla el período Medio o Tiwanaku con las fases Quitor
y Coyo y cuando declina la influencia de Tiwanaku se inicia el período
Intermedio Tardío o de Desarrollo Regional (1.000 d.C. – 1.470 d.C.) con las fases Yaye y Solor, el cual culmina
con el dominio Inka (1.471 d.C. - 1.532 d.C.) en la región (Berenguer et al. 1986; Llagostera et al. 1988; Tarragó, 1989). En la
Figura 1 se presentan los sitios en los cuales se obtuvieron las muestras
empleadas en el presente trabajo.
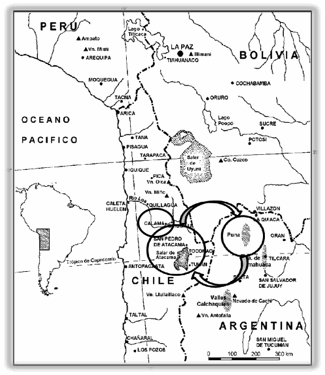
Figura
1.
Ubicación de las subáreas Puna de Jujuy, San Pedro de Atacama y Calama
Figure
1.
Location of the sub-areas Puna de Jujuy, San Pedro de Atacama and Calama
A 101 km al NO de SPA,
a orillas del río Loa, se
encuentra el sitio Chunchurí en la localidad de Calama, datado en 1.390 d.C.
(sitio Dupont-1) (Nuñez 1965, 1968 y 1976; Nuñez et al. 2003; Núñez y Pimentel 2008) y corresponde al Período Tardío
pre Incaico. Parte del material cultural de este sitio obtenido por Max Uhle en
1912 fue descripto por Durán et al
(2000) y los restos bioantropológicos por Cocilovo et al. (2016). Desde SPA, a 427 km (promedio) en dirección SO en
territorio argentino, se encuentran un conjunto de sitios de la PJ del Período
Tardío (Agua Caliente, Casabindo, Doncellas, Queta, Río Negro, Sorcuyo y
Tablada de Abra Pampa) fechados entre 1.029 - 1.208 d.C. (±2σ) y 1.400 - 1.497 d.C. (±2σ) (Fuchs, 2014; Fuchs y
Varela, 2013). El trayecto entre Calama y Puna de Jujuy es de 528 km (promedio).
El estudio de la estructura de la
población se basó en el análisis de 1.107 individuos de las subáreas: PJ (323),
CAL (161) y SPA (623). La muestra está integrada por individuos de ambos sexos,
de edades adulto, maduro y senil, con deformación tabular erecta y oblicua,
circular y no deformados. La determinación del sexo, de
la edad y de la deformación artificial del cráneo se realizó de acuerdo con
Genovés (1962), Dembo e Imbelloni (1938) y Buikstra y Ubelaker (1994). Se
midieron 31 variables métricas (Tabla 1) de acuerdo con la Convención
Internacional de Mónaco de 1906 (Comas, 1966), Wilder (1920) y Bass (1981)
empleando instrumentos originales de Siber y Hegner (Suiza).
|
Variables |
||
|
Longitud máxima |
Diámetro nasio basilar |
Longitud maxilo alveolar |
|
Anchura máxima |
Diámetro alvéolo basilar |
Longitud del paladar |
|
Altura basilo bregmática |
Altura nasio alveolar |
Anchura del paladar |
|
Altura porio bregmática |
Altura de la naríz |
Altura órbito alveolar |
|
Diámetro frontal mínimo |
Anchura de la naríz |
Longitud foramen magnun |
|
Diámetro frontal máximo |
Altura del pómulo |
Anchura foramen magnun |
|
Anchura fronto malar |
Anchura biorbitaria |
Curva nasion bregma |
|
Anchura bicigomática |
Anchura de la órbita |
Curva bregma lambda |
|
Anchura bimaxilar máxima |
Altura de la órbita |
Curva lambda opistion |
|
Anchura biastérica |
Anchura maxilo alveolar |
Curva horizontal |
|
Anchura biauricular |
|
|
Tabla 1. Variables empleadas en el presente trabajo
Table 1. Variables used in the present study
Las diferencias entre subregiones se evaluaron mediante técnicas
estadísticas de análisis multivariado: MANOVA y Análisis Discriminante (Seber,
1984). El estudio incluyó la evaluación de la clasificación de los individuos al
propio grupo o a uno distinto junto con la probabilidad condicional asociada según
la Regla de Bayes (Montes
Suay, 2007). El resultado de este último cálculo permitió determinar: 1) la
homogeneidad de las muestras de cada subregión y 2) analizar los casos
asignados a otros grupos.
La divergencia genética entre las poblaciones fue evaluada mediante
valores Fst para rasgos cuantitativos de acuerdo con Relethford (1994, 1996,
2001 y 2002), Relethford y Blangero (1990), Relethford y Harpending (1994) y Relethford et al. (1997). El estadístico Fst mide la magnitud de la
diferenciación genética y la semejanza entre los grupos a partir de la relación
entre la varianza entre grupos y la varianza promedio dentro de grupos (Varela
y Cocilovo, 2000, 2002). Esta relación varía de acuerdo con el efecto de la
deriva genética y de la migración en ausencia de selección y de mutación,
aunque Fst es relativamente insensible al efecto de estos factores (Slatkin
1994). El valor de Fst se calculó para caracteres continuos con una
heredabilidad de 0,55, entre subáreas y
entre sexos por subáreas para evaluar el comportamiento reproductivo de los
individuos a nivel local y regional. Además este cálculo permite inferir la
magnitud del flujo génico local o proveniente de una fuente externa a la región
estudiada.
Resultados
La distribución del sexo (Tabla 2) refleja una
mayoría de individuos masculinos (58 %). En cada subárea la proporción de ambos
sexos varía, es semejante en SPA y los hombres son más frecuente en PJ (72 %) y
CAL (65 %), siendo en general las diferencias estadísticamente significativas
(Pearson Chi-square: 32,59, gl=2, p<0,01).
|
Subarea\Sexo |
Masculino |
Femenino |
Total |
|
San Pedro de Atacama |
319 |
304 |
623 |
|
% |
0,512 |
0,488 |
|
|
Calama (Chunchurí) |
116 |
45 |
161 |
|
% |
0,720 |
0,280 |
|
|
Puna de Jujuy |
212 |
111 |
323 |
|
% |
0,656 |
0,344 |
|
|
Total |
647 |
460 |
1107 |
Tabla 2. Denominación y composición de las muestras
Table 2. Denomination and composition
of samples
El análisis de las diferencias fenotípicas entre subregiones (CAL, SPA y
PJ) reveló resultados significativos (Tabla 3). La diferencia global fue
evaluada por la prueba de Wilks' Lambda: 0,26315 approx. F(62 y 2148gl)
= 32,892, p < 0,01. Estos resultados se traducen en la Figura 2 en donde se
muestra la distribución de los individuos de cada grupo según las dos primeras
coordenadas discriminantes que acumulan el 100 % de la variación. En dicha
figura se observan las áreas de la
distribución y la proporción del solapamiento entre las mismas. Además los
valores D2 (Tabla 3) se correlacionan con las distancias espaciales
entre las muestras de cada subárea (Coeficiente Correlación r = 0,94).
|
Subregiones |
Calama (Chunchurí) |
San Pedro de Atacama |
Puna de Jujuy |
|
Calama
(Chunchurí) |
0,00 |
5,04 |
9,47 |
|
San
Pedro de Atacama |
5,04 |
0,00 |
7,09 |
|
Puna
de Jujuy |
9,47 |
7,09 |
0,00 |
Tabla 3. Valores D2, diferencias entre
fenotipos medios de subáreas. Para todos los casos
p=0; Wilks' Lambda: 0,26315 approx. F (62,2148)=32,892 p<0,0000
Table
3. D2 values,
differences between mean phenotypes of subareas. For all cases p = 0; Wilks'
Lambda: 0.26315 approx. F (62.2148) = 32.892 p <0.0000
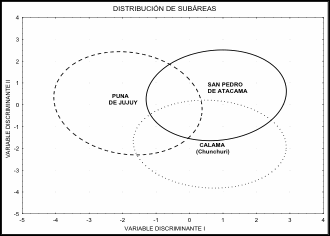
Figura 2. Distribución de las subregiones por las
coordenadas discriminantes (Wilks' Lambda: 0,26315 aproximación F (62 y 2148 gl)
= 32,892, p < 0,01) Variación
explicada 100 %.
Figure
2. Distribution of the
subregions by the discriminant coordinates (Wilks' Lambda: 0.26315 F
approximation (62 and 2148 df) = 32.892, p < 0.01). Variation explained 100 %.
En la prueba de la clasificación se comprobó que el 87 % de los
individuos fue asignado al grupo original (Tabla 4). En SPA se observó la mayor
proporción de casos clasificados al propio grupo y la menor para CAL. El 13 %
fue clasificado en un grupo distinto del original con 68 % masculinos y 32 %
femeninos. La distribución entre sitios refleja igualmente el espaciamiento
geográfico entre ellos (Tabla 4). Esto último, se destaca también en la tabla 5
en la cual se presentan las proporciones de individuos reclasificados a un
grupo distinto del original y el valor medio de la probabilidad para cada
grupo. Por ejemplo, desde CAL a SPA se registró el 29 % (43/150) de individuos
(![]() = 0,70 de probabilidad) y a PJ con
el 6 % (9/150) de individuos (
= 0,70 de probabilidad) y a PJ con
el 6 % (9/150) de individuos (![]() = 0,57 de probabilidad), desde SPA
la proporción de individuos asignados a CAL fue de 13 % (19/150) y de 23 % (34/150)
a PJ, con valores medios de probabilidad de 0,69 y 0,72 respectivamente y desde
PJ el 24 % de los casos (36/150) a SPA (
= 0,57 de probabilidad), desde SPA
la proporción de individuos asignados a CAL fue de 13 % (19/150) y de 23 % (34/150)
a PJ, con valores medios de probabilidad de 0,69 y 0,72 respectivamente y desde
PJ el 24 % de los casos (36/150) a SPA (![]() = 0,67 de probabilidad) y el 6 % (9/150)
a CAL (
= 0,67 de probabilidad) y el 6 % (9/150)
a CAL (![]() = 0,55 de probabilidad).
= 0,55 de probabilidad).
|
Subregiones |
Porcentaje Correcto |
Calama (Chunchurí) p=,14544 |
San Pedro de Atacama p=,56278 |
Puna de Jujuy p=,29178 |
|
Calama (Chunchurí) |
67,70 |
109 |
43 |
9 |
|
San Pedro de Atacama |
91,49 |
19 |
570 |
34 |
|
Puna de Jujuy |
86,07 |
9 |
36 |
278 |
|
Total |
86,45 |
137 |
649 |
321 |
Tabla
4.
Análisis de la reclasificación de los individuos y probabilidad de pertenecer
al propio grupo. Total de
individuos reclasificados: 150 (14 %)
Table 4. Analysis of the reclassification of the
individuals and probability of belonging to the group. Total reclassified
individuals: 150 (14 %),
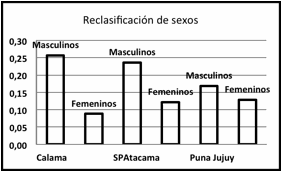
En la Figura 3 se presenta el análisis de agrupamiento (método de Ward)
entre los sexos en base a los valores D2 en donde se destaca la
consistencia de los fenotipos de ambos sexos para cada subregión (Wilks'
Lambda: 0,11427 approx. F(155,5309) = 18,847 p < 0,01). En la
Tabla 6 se presentan los valores D2 para hombres y mujeres entre
subregiones y las probabilidades de las pruebas de significación. Se destaca la
semejanza entre los valores D2 de masculinos y femeninos entre
subáreas y el valor creciente de las diferencias entre cada sexo con relación a
la distancia geográfica.
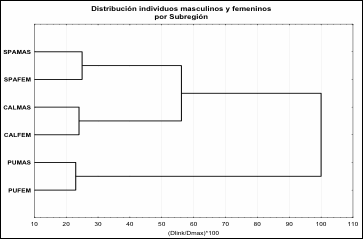
Figura
3.
Asociación de los sexos por subregión en base a valores D2 Método de
Ward. SPAMAS (San Pedro de Atacama masculinos), SPAFEM (San pedro de Atacama
femeninos), CALMAS (Calama masculinos), CALFEM (Calama femeninos), PUMAS (Puna
de Jujuy masculinos) y PUFEM (Puna de Jujuy femeninos).
Figure 3. Association of the sexes by subregion
based on D2 values Ward's method. SPAMAS (San Pedro de Atacama male), SPAFEM (San
pedro de Atacama female), CALMAS (male Calama), CALFEM (female Calama), PUMAS
(male Jujuy Puna) and PUFEM (female Jujuy Puna).
Es interesante señalar que la reasignación de individuos en una u otra
subregión sigue un patrón definido (Figura 4 a 6). Por ejemplo, desde SPA a CAL
y PJ se destaca la contribución de
sitios del Período Medio y Tardío como Coyo Oriental, Quitor 5, Quitor 6, Yaye
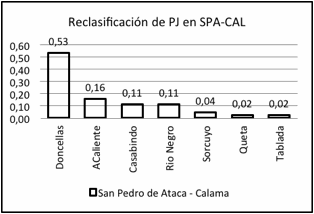
y Quitor 1 y, desde PJ a SPA y CAL desde los sitios Doncellas, Agua Caliente,
Casabindo y Río Negro correspondientes al Período Intermedio Tardío. También
sigue un patrón definido la reclasificación de los individuos de cada subregión
por sexo (Figura 6) en una subregión distinta de la original: desde CAL la
proporción de masculinos asignados a otros grupos es decreciente y la de
femeninos aumenta moderadamente hasta PJ.
|
Subregiones |
Índices |
Calama
(Chunchurí) |
San Pedro de
Atacama |
Puna de Jujuy |
|
Calama
(Chunchurí) |
% |
------ |
29 |
6 |
|
|
|
0,7 |
0,57 |
|
|
San Pedro de
Atacama |
% |
13 |
------ |
23 |
|
|
0,69 |
0,72 |
||
|
Puna de Jujuy |
% |
6 |
24 |
------ |
|
|
0,55 |
0,67 |
Tabla
5.
Proporción y probabilidad media de los individuos reclasificados entre subregiones.
Total de individuos
reclasificados: 150 (14 %).
Table 5. Proportion and average probability of
reclassified individuals between subregions.
Total reclassified
individuals: 150 (14 %).
La estructura de la población a nivel regional (Figuras 7 y
8) evaluada a partir del estadístico Fst = 0,063 ± 0,002,
indica un proceso de divergencia determinado por un balance distinto del
esperado por el modelo entre el efecto de la deriva y de la migración. Se
destaca PJ por presentar una varianza fenotípica mayor que la esperada a
diferencia de SPA y CAL cuyas varianzas son menores, posiblemente por una mayor
interacción entre localidades de la propia región, mientras que PJ refleja la
influencia de variabilidad genética de un rango territorial mayor.
|
Subregión |
Calama (Chunchuri) |
San Pedro de Atacama |
Puna de Jujuy |
||||
|
Sexo |
Masculinos |
Femeninos |
Masculinos |
Femeninos |
Masculinos |
Femeninos |
|
|
Calama (Chunchuri) |
Masculinos |
0,00 |
4,64 |
5,81 |
7,06 |
9,30 |
13,26 |
|
Femeninos |
4,64 |
0,00 |
11,87 |
6,41 |
13,57 |
12,80 |
|
|
San Pedro de Atacama |
Masculinos |
5,81 |
11,87 |
0,00 |
4,82 |
8,40 |
13,00 |
|
Femeninos |
7,06 |
6,41 |
4,82 |
0,00 |
9,24 |
7,53 |
|
|
Puna de Jujuy |
Masculinos |
9,30 |
13,57 |
8,40 |
9,24 |
0,00 |
4,44 |
|
Femeninos |
13,26 |
12,80 |
13,00 |
7,53 |
4,44 |
0,00 |
|
Tabla 6. Valores D2, diferencias entre individuos
de ambos sexos por subarea. Diferencia global: Wilks' Lambda: 0,11427 approx. F(155,5309)
gl = 18,847 p=0. Diferencia entre pares de comparaciones (entre sexos y
entre subregiones) p=0.
Table 6. D2 values, differences between
individuals of both sexes by subarea. Overall Difference: Wilks' Lambda: 0.11427
approx. F (155.5309 df) = 18.847 p = 0. Difference between pairs of
comparisons (between sexes and between subregions) p = 0.
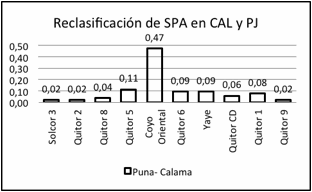
Figura
4.
Reclasificación de individuos de San Pedro de Atacama en Calama y Puna de
Jujuy. Se especifican las proporciones desde cada sitio de San Pedro de
Atacama.
Figure 4. Reclassification of individuals from San
Pedro de Atacama in Calama and Puna de Jujuy. The proportions are specified
from each site of San Pedro de Atacama.
Figura
5.
Reclasificación de individuos de Puna de Jujuy en San Pedro de Atacama y
Calama. Se especifican las proporciones de cada sitio de Puna de Jujuy.
Figure 5. Reclassification of individuals from Puna
de Jujuy in San Pedro de Atacama and Calama. The proportions of each site of
Puna de Jujuy are specified.
Figura
6.
Proporción de individuos reclasificados por sexo desde la subregión de origen
en una subregión distinta de la original.
Figure 6. Proportion of individuals reclassified by
sex from the subregion of origin in a sub-region other than the original one.
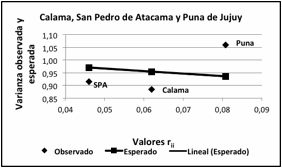
Figura 7. Varianza fenotípica observada y esperada para
las subregiones Calama, San Pedro de Atacama y Puna de Jujuy. Valores rii
distancia al centroide de la distribución. Valor Fst = 0,063 ± 0,002.
Figure
7. Observed and expected
phenotypic variance of the Calama, San Pedro de Atacama and Puna de Jujuy
subregions. Values rii distance to the centroid of the distribution. Fst value = 0.063 ±
0.002.
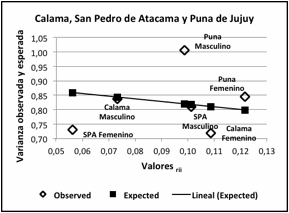
Figura 8. Varianza
fenotípica observada y esperada de las subregiones Calama, San Pedro de Atacama
y Puna de Jujuy para sexo masculino y femenino. Valores rii distancia al
centroide de la distribución. Valor Fst = 0,0931 ± 0,003.
Figure 8. Observed and expected phenotypic variance of the Calama, San
Pedro de Atacama and Puna of Jujuy subregions for male and female. Values rii
distance to the centroid of the distribution. Fst value = 0.0931 ± 0.003.
Discusión
Las diferencias fenotípicas entre subáreas (Tabla 3) es una información
que amplía los resultados de experiencias anteriores (Varela et al., 1999 y 2008) y confirma la
existencia desde el Período Medio y Tardío regional de historias de vida
diferentes afectando a la mayoría de los habitantes de uno y otro lado de la
Cordillera de los Andes. Las diferencias de las
distribuciones de los sexos entre subáreas comprobada en este trabajo (Tabla
2) es un hecho relevante advertido anteriormente al
estudiar las relaciones entre SPA y PJ (Fuchs et al., 2016a; Cocilovo et al.,
2017), tal vez marcando pautas culturales y laborales diferentes relacionadas con
el tránsito migratorio y caravanero.
La influencia de la distancia entre las subáreas fue un factor
importante en la diferenciación global y particular entre ambos sexos (Tablas 3
y 6). Este efecto se manifestó también en la reclasificación de los individuos
entre las subáreas (Tabla 4) y en los valores de probabilidad asociados con las
asignaciones (Tabla 5). El nivel de interacción regional, sin omitir momentos
más tempranos, fue más intenso en los períodos Medio y Tardío e involucró más a
individuos masculinos (68 %) que femeninos (32 %) lo cual se expresa en la
distribución por sexo de la Figura 6. Estas observaciones también se obtuvieron
en un estudio previo entre PJ y SPA (Cocilovo et al., 2017).
La reasignación de casos por el análisis discriminante es consistente
desde el punto de vista cronológico pues la frecuencia se concentra entre los
períodos Medio y Tardío. Para SPA (Figura 4), considerando las fechas más
tardías: Quitor 1: 956 ± 44 a.P. (cal d.C. 1.032 - 1.211), Quitor 5: 1.164 ± 44
a.P. (cal 782 - 1.019 d.C.), Quitor 6: 1.050 ± 40 a.P. (cal 982 - 1.150 d.C.),
Yaye 1: 920 ± 40 a.P. (cal 1.042 - 1.260 d.C.)
(Hube et al., 2011). Y para el
sitio Chunchurí (CAL), datado en 1.390 d.C. (sitio Dupont-1) (Nuñez, 1965, 1968
y 1976; Nuñez et al. 2003; Núñez y
Pimentel, 2008). La muestra de PJ posee fechas radiocarbónicas, que cubren el
intervalo de 964 ± 45 a.P. (cal d.C. 1.029 - 1.208 ± 2σ) para el Pucará de
Rinconada y 501 ± 42 a.P. (cal d.C. 1.400 -1.497 ± 2σ) para Casabindo (Fuchs y Varela, 2013) incluyendo los sitios
de la Figura 5.
En la composición de PJ el valor de la varianza fenotípica observada con
respecto a la esperada (Figura 7 y 8) refleja el efecto de un rango mayor de
interacción genética que involucra a ambos sexos pero en mayor proporción al
masculino con relación a SPA y CAL, mientras que para el componente femenino
de estas subáreas la varianza fenotípica
es mucho menor. En efecto, considerando solamente las tres subregiones la
divergencia medida por el estadístico Fst es 6,3 %, y al evaluar ambos sexos
por subregión se eleva al 9,3 %. El incremento de ambos estadísticos es notable
comparado con los valores obtenidos en un estudio anterior entre SPA y PJ: Fst =
2,4 % y Fst = 6 % entre los sexos de ambas poblaciones (Cocilovo et al. 2017). Se destaca el efecto de la
distancia en la divergencia genética a nivel regional indicando la extinción
gradual del parentesco entre subáreas.
Es interesante comparar estos resultados con los obtenidos en otros estudios
que revelan variaciones relacionadas con la estructura temporal, espacial y
sexual. En el ayllu de Quitor de SPA se estimó una divergencia promedio de 5 %
entre los Períodos Temprano, Medio y Tardío con un valor global Fst mayor en
mujeres (7,6 %) que en varones (6,4 %) (Varela y Cocilovo, 2009) lo cual dejó
planteado para este sitio el tema de la mayor diversidad femenina por un menor
tamaño efectivo con relación a los hombres. Precisamente, en la experiencia
actual (Figura 8) los grupos femeninos de SPA y CAL mostraron una varianza
fenotípica menor que la esperada por un mayor aislamiento y la interacción en
un ámbito territorial menor. En cambio, se destaca la desviación de este
equilibrio de los individuos masculinos de PJ indicando mayor diversidad
genética y una interacción regional mucho más amplia. Esta observación repite
una medición anterior realizada a nivel del Área Andina Centro Sur (Varela et al. 2012).
En una muestra mayor de SPA incluyendo una decena de sitios, el valor
Fst fue muy similar al anterior (5 %), y se comprobó la variación de este
indicador en el tiempo: Período Temprano Fst = 0,114 ± 0,0163, Período Medio
Fst = 0,005 ± 0,004 y Período Tardío Fst = 0,026 ± 0,009 (Varela y Cocilovo,
2011). Estos valores de Fst marcan la historia de esta subregión relacionada
con el poblamiento original y con el aporte de variabilidad genética externa. La
divergencia fenotípica estimada entre SPA y PJ (2,4 %) refleja la conservación
de la mayor variación genética dentro de cada subárea alcanzando el valor de 8 %
cuando se consideran los sitios de cada
subregión (Cocilovo et al. 2017). Por
otra parte, en la experiencia actual, al incluir Calama en el análisis, la divergencia
entre subáreas (Fst = 6,3 %) se incrementó un 45 % por el aporte de mayor
variabilidad contenida en esta última subárea.
También es importante destacar la concordancia entre las mediciones de
variabilidad fenotípica (Fst) obtenidas para fases tardías del desarrollo
regional. Por ejemplo en PJ con una composición del Período Intermedio Tardío,
se obtuvo el valor de 0,025 (Fuchs et al.
2016a), semejante al estimado para el período final del desarrollo de SPA de 0,026
(Varela y Cocilovo, 2011) y al valor global determinado entre PJ y SPA de 0,024
(Cocilovo et al. 2017). Estos valores
pueden ser explicados por el aumento de la población regional coincidente con el
incremento de la señal en 4700 a.P. cal
(Muscio y Lopez, 2016) y una mayor interacción entre distintas subáreas que
estimuló la migración y el intercambio económico. Por ejemplo, se demostraron
relaciones basadas en la ausencia de diferencias fenotípicas significativas
entre sitios de PJ y SPA: Doncellas, Queta y Río Negro con Quitor CD y Yaye
(Fuchs et al. 2016b) y una mayor
interacción regional de los sitios: Larache, Quitor CDB y CDN, Solcor 3, Agua
Caliente y Casabindo (Cocilovo et al.,
2017). Además, esta relación se proyecta territorialmente por la ausencia de diferencias
significativas basadas en estimaciones de D2 entre PJ y sitios de la
Quebrada de Humahuaca como Alfarcito, Angosto Chico, La Huerta y Pukará de
Humahuaca (Varela et al. 2017). Tal
vez, este nivel de interacción se relacione con la conservación de una mayor
frecuencia del linaje A2 entre SPA y PJ, que también se manifiesta en la
Quebrada de Humahuaca (de Saint Pierre et
al. 2012; Mendisco et al. 2014;
Postillone et al., 2017).
En síntesis, se comprobaron diferencias fenotípicas significativas a
nivel regional entre Puna de Jujuy, San Pedro de Atacama y Calama (Chunchuri).
Dichas diferencias se correlacionan con las distancias espaciales entre
subregiones. El dimorfismo sexual se expresa con valores semejantes en cada
subárea y las diferencias entre masculinos y femeninos de cada subárea también siguen
un patrón espacial. La proporción de individuos reclasificados en una subregión
distinta de la original fue de 13 %, involucrando un mayor porcentaje de
hombres que de mujeres. La proporción reasignada y la probabilidad media varían
de acuerdo con la distancia entre subregiones, entre los períodos Medio y Tardío.
Esta evidencia tal vez se relaciona con actividades laborales y culturales
diferentes desempeñados por ambos sexos. La estructura de la población regional
estuvo determinada por un balance desigual entre la deriva y la migración involucrando
ambos sexos. Las diferencias fenotípicas comprobadas explican a nivel regional el
proceso evolutivo por el cual se diferenciaron las poblaciones del Norte de
Chile de las del Noroeste Argentino. Los valores Fst estimados son el resultado
del equilibrio entre el tamaño de las poblaciones locales y el aporte de
novedades genéticas externas. La información sobre DNAmt disponible refleja la
integración de las distintas sociedades con linajes locales y compartidos a
nivel regional. Sería sumamente interesante completar el presente estudio con
otras localidades de la región y particularmente con aquellas que permitan
completar el trayecto hasta la costa Pacífica.
Agradecimientos. Agradecemos la
colaboración brindada por las autoridades y el personal de los museos en donde se
encuentran depositadas las colecciones osteológicas empleadas en este trabajo:
Museo Nacional de Historia Natural en Santiago (Chile), Instituto de
Investigaciones Arqueológicas R.P. Gustavo Le Paige, S.J. en San Pedro de
Atacama, Museo Etnográfico "Juan Bautista Ambrosetti" de la
Universidad Nacional de Buenos Aires y Museo de Ciencias Naturales de La Plata.
El presente trabajo se realizó con los siguiente fondos: Consejo Nacional de
Investigaciones Científicas y Técnicas (CONICET, PIP 2405/08), Argentina;
Secretaría de Ciencia y Técnica-Universidad Nacional de Río Cuarto
(SeCyT-UNRC), Argentina.
Bibliografía
Aldenderfer, M.S. y Flores Blanco L., 2011 Reflexiones para Avanzar en los Estudios del Período Arcaico en los Andes Centro-Sur Chungara. Revista De Antropología Chilena 43 (Número Especial), 531-550.
Barbujani, G., 1987, Autocorrelation of Gene Frequencies Under Isolation by Distance. Genetics 117, 777-782.
Bass, W.M., 1981, Human Osteology: A laboratory and field manual of
the human skeleton. University
of Missouri Press, Springfield.
Berenguer, J.A., Deza, A.R. y Llagostera, A., 1986 La secuencia de Myriam Tarragó para San
Pedro de Atacama: Un test por termoluminiscencia. Revista Chilena de Antropología 5,17-54
Buikstra,
J.E. y Ubelaker, D., 1994, Standards for data collection from human skeletal
remains. Fayetteville, Arkansas: Arkansas archeological survey research series
nº 44.
Cocilovo, J.A., 1981, Estudio sobre discriminación y clasificación de
poblaciones prehispánicas del N. O. Argentino. Publicación Ocasional 36, 3-60
Museo Nacional de Historia Natural, Santiago de Chile.
Cocilovo, J.A. y Di Rienzo, J.A., 1984-1985, Un Modelo Biológico para el
Estudio del Poblamiento Prehispánico del Territorio Argentino. Correlación
Fenético-Espacial. Relaciones de la Sociedad Argentina de Antropología
16,119-135.
Cocilovo, J.A
y Rothhammer, F., 1990, Paleopopulation biology of southern Andes: craniofacial
chronological and geographical differentiation. Homo
411,16-31
Cocilovo, J.A. y Varela, H.H., 1999, Variación Morfológica, Estructura
Canónica y Bioensayo de Parentesco en Poblaciones Prehistóricas del Norte de
Chile. Chungará (Arica) 30 (1),75-85.
Cocilovo, J.A., Varela, H.H. y Fuchs, M.L., 2017, Movilidad, Tráfico Sur
Andino y Diferenciación Genética entre
San Pedro de Atacama y la Puna De Jujuy. Manuscrito.
Cocilovo, J.A., Varela, H.H. y Quevedo, S., 1995, La deformación
artificial del cráneo en la población prehistórica de San Pedro de Atacama,
Chile. Chungara 27(2),117-124.
Cocilovo, J.A., Varela, H.H. y Valdano, S.G., 2016, La Población
Prehistórica de Calama Chunchuri Norte de Chile. Composición y Estructura.
Boletín del Museo Nacional de Historia Natural Chile 65, 291-306.
Cocilovo, J.A., Zavattieri, M.V. y Costa Junqueira, M.A., 1994, Biología
del grupo prehistórico de Coyo Oriental (San Pedro de Atacama, Noprte de
Chile): I Dimorfismo sexual y variación etaria, Estudios Atacameños 11,121-134.
Comas, J. 1966. Manual d e antropología física, Instituto de
Investigaciones Históricas. México: Universidad Nacional Autónoma de México.
Costa, M.A. y Llagostera, A., 2014, Leishmaniasis en Coyo Oriente.
Migrantes trasandinos en San Pedro de Atacama. Estudios Atacameños Arqueología
y Antropología Surandinas 47, 5-18.
Costa, MA.,
Matheson, C., Iachetta, L., Llagostera,
A. y Appenzeller, O., 2009, Ancient Leishmaniasis in a Highland Desert of
Northern Chile. PLoS ONE, 4(9) e6983.
doi:10.1371/journal.pone.0006983.
De Nigris, M. 2009. Arqueología, Minería y Metalurgia en la localidad de
Cobres y sus alrededores (Períodos Prehispánico y Colonial). Tesis para optar
por el título de Antropólogo, Escuela de Antropología, Facultad de Humanidades,
Universidad Nacional de Salta. Salta.
De Saint
Pierre, M., C. Bravi, Motti, J.M.B.,
Fuku, N., Tanaka, M., Llop, E., Bonatto, S. y Moraga, M., 2012, An Alternative
Model for the Early Peopling of Southern South America Revealed by Analyses of
Three Mitochondrial DNA Haplogroups, PLoS ONE 7(9): e43486.
Dembo, A. e Imbelloni, J.. 1938 Deformaciones intencionales del cuerpo
humano. Humanior, Sección A 3. Buenos Aires: Imprenta Luis L. Gotelli.
Durán E., Kangiser, M.F. y Acevedo, N., 2000, Colección Max Uhle:
Expedición a Calama 1912 Publicación Ocacional del Museo Nacional de Historia
Natural Chile 56, 5-49.
Fuchs, M.L., 2014. Caracterización bioestructural de la población
antigua de la Puna de Jujuy (Argentina). Tesis doctoral inédita, Facultad de
Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río
Cuarto, Argentina.
Fuchs, M.L. y Varela, H. 2013, Fechados radiocarbónicos de Colecciones
osteológicas de la Puna de Jujuy, Argentina. Relaciones de la Sociedad Argentina
de Antropología 38, 553-558
Fuchs, M.L.,
Varela, H.H. y Cocilovo, J.A., 2016a, Kinship and Phenotypic Divergence in the
Ancient Population of the Puna Plateau of Northwestern Argentina. Advances in Anthropology 6, 1-10.
Fuchs, M.L., Varela, H.H., Cocilovo, J.A., 2016b, Relaciones biológicas
entre la Puna de Jujuy Argentina y San Pedro de Atacama Chile. Antropo 36,
15-27. www.didac.ehu.es/antropo
Genovés, S., 1962, Introducción al Diagnóstico de la Edad y del Sexo en
Restos Óseos Prehistóricos. Publicación del Instituto de Historia, México.
Hartl, D.L. y Clark, A.G., 1997, Principles of population
genetics. 3nd ed. By. Sunderland, M.A, Sinauer Associates, Inc.
Hubbe, H., Oviedo, M. y Torres-Rouff, C., 2011, Estado de conservación y contextualización
cronológica de la Colección Osteológica “Gustavo Le Paige”. Estudios Atacameños
Arqueología y Antropología Surandinas 41,29-44
Knudson,
K.J., Pestle, W.J., Torres-Rouff, C. and Pimentel, G., 2012, Assessing The Life
History of an Andean Traveller through Biogeochemistry: Stable and Radiogenic
Isotope Analyses of Archaeological Human Remains from Northern Chile. Int. J. Osteoarchaeol. 22, 435–451.
Llagostera, A. 1995. El componente cultural Aguada en San Pedro de
Atacama. Boletín del Museo Chileno de Arte Precolombino, 6, 9-34.
Llagostera, A., Torres, C.M. y Costa, M.A., 1988, Complejo psicotrópico
de Solcor-3 (San Pedro de Atacama). Estudios Atacameños 9,61-98.
Mendisco, F.,
Keyser, Ch., Seldes, V., Rivolta, C., Mercolli, P., Cruz, P., Nielsen, A. E.,
Crubezy, E. and Ludes, B., 2014, Genetic Diversity of a Late Prehispanic Group
of the Quebrada de Humahuaca, Northwestern Argentina. Annals of Human Genetics,
78(5), 367-380.
Molina Otárola, R., 2011, Los otros arrieros de los valles La Puna y el
Desierto de Atacama. Chungara Revista de Antropología Chilena 43 2 , 177-187
Montenegro, M., y Ruiz, M. 2007. Tránsito y paisaje en la puna de Jujuy
durante los desarrollos regionales: una aproximación iconográfica. Cuadernos de
la Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy,
(32), 167-185.
Montes Suay, F., 2007 Introducción a la Probabilidad. Departamento de
Estadística e Investigación Operativa, Universitad de Valéncia, España.
Morton, N.E.,
1982 Outline of Genetic Epidemiology, S. Karger, New York.
Muscio, H.
J., y López, G. E. 2016. Radiocarbon dates and anthropogenic signal in the
South-Central Andes (12,500–600 cal. years BP). Journal of Archaeological
Science, 65, 93-102.
Nielsen, A.E., 2002, Asentamientos, conflicto y cambio social en el
altiplano de Lípez (Potosí). Revista Española de Antropología Americana 32,
179-205.
Nielsen, A.E. 2011. El tráfico de caravanas entre Lípez y Atacama visto
desde la Cordillera Occidental. En Ruta: Arqueología, Historia y Etnografía del
Tráfico Sur Andino, editado por L. Núñez A, y AE Nielsen, 83-110.
Núñez, L. 1965. Desarrollo cultural prehispánico del norte de Chile.
Estudios Arqueológicos, 1, 37-115.
Nuñez, L., 1968, Subárea Loa-Costa Chilena desde Copiapó a Pisagua.
Actas 32 Congreso Internacional de Americanistas, pp. 145-182, Buenos Aires.
Nuñez, L., 1976, Registro regional de fechas radiocarbónicas en el Norte
de Chile. Estudios Atacameños 4,74-123.
Núñez A.L., Agüero P.C., Cases
C.B. y De Souza H. P., 2003, El campamento minero Chuquicamata-2 y la
explotación cuprífera prehispánica en el Desierto de Atacama. Estudios
Atacameños 25,27-34
Núñez L. y Dillehay, T., 1995, Movilidad giratoria armonía social y
desarrollo en los Andes Meridionales: Patrones de tráfico e interacción
económica. Universidad Católica del Norte Antofagasta 2o ed. 190 pp.
Núñez, L. y Pimentel, G., 2008, Informe prospección arqueológica
proyecto exploración geológica de óxidos y sulfuros, División Codelco Norte,
Chile.
Postillone, M.B., Fuchs, M.L., Crespo, C.M., Russo, M.G., Varela, H.H.,
Carnese, F.R., Avena, S.A. y Dejean, C.B., 2017, Linajes Maternos en Muestras
Antiguas de la Puna Jujeña. Comparación con Estudios de la Región Centro-Sur
Andina. Revista Argentina de Antropología Biológica, 19(1).
Relethford,
J.H., 1994, Craniometric variation among modern human populations. Am. J. Phys.
Anthropol. 95,53–62.
Relethford,
J.H., 1996, Genetic drift can obscure population history: problem and solution.
Hum Biol 68, 29–44.
Relethford,
J.H., 2001, Global analysis of regional differences in craniometric diversity
and population substructure. Hum Biol 73, 629–636.
Relethford,
J.H., 2002, Apportionment of Global Human Genetic Diversity Based on
Craniometrics and Skin Color. American Journal of Physical Anthropology 118,
393-398.
Relethford,
J.H., and Blangero, J., 1990. Detection of differential gene flow patterns of
quantitative variation. Hum Biol 62,5–25.
Relethford,
J.H. and Harpending, H.C., 1994, Craniometric variation, genetic theory, and
modern human origins. Am J. Phys. Anthropol. 95,249–270.
Relethford,
J.H., Crawford M.H., Blangero, J., 1997, Genetic drift and gene flow in
post-famine Ireland. Hum Biol 69,443-465.
Seber,
G.A.F., 1984, Multivariate observation. Ed. John Wiley and Son, Nueva York.
Slatkin, M.,
1994, Gene flow and population structure. En
Ecological Genetics, Ed. por L. Real. Princeton.
Tarragó, M., 1977, Relaciones prehispánicas entre San Pedro de Atacama,
Norte de Chile y regiones aledañas: la Quebrada de Humahuaca. Estudios Atacameños
5, 50-63.
Tarragó, M., 1984, La historia de los pueblos circumpuneños en relación
con el altiplano y los Andes Meridionales. Estudios Atacameños 7,93-104.
Tarragó, M.N., 1989, Contribución al conocimiento arqueológico de las
poblaciones de los oasis de San Pedro de Atacama en relación con los otros
pueblos puneños, en especial, el sector septentrional del valle Calchaquí.
Tesis para optar el título de Doctor en Historia, Especialidad
Arqueología, Facultad de Humanidades y
Artes, Universidad Nacional de Rosario, Argentina.
Torres-Rouff,
Ch., Knudson, K.J., Pestle, W.J. and Stovel, E.M., 2015 Tiwanaku Influence and
Social Inequality: A Bioarchaeological, Biogeochemical, and Contextual Analysis
of the Larache Cemetery, San Pedro de Atacama, Northern Chile. American Journal
of Physical Anthropology 158,592–606 .
Varela, H.H. y Cocilovo, J.A., 1996, Los componentes de la variación
intramuestral en la población prehistórica de San Pedro de Atacama, Chile.
Revista Argentina de Antropología Biológica 1(1), 219-231.
Varela, H.H.
y Cocilovo, J.A., 2000, Structure of the Prehistoric Population of San Pedro de
Atacama. Current Anthropology, 41(1), 125-132 The University of Chicago Press,
Wenner-Gren Foundation for Anthropological Research.
Varela, H.H. y Cocilovo, J.A., 2002. Genetic Drift and Gene
Flow in a Prehistoric Population of the Azapa Valley and Coast, Chile. American
Jorunal of Physical Anthropology 118, 259-267.
Varela H.H. y Cocilovo, J.A., 2009. Microevolución en San Pedro de
Atacama (Norte de Chile): el cementerio de Quitor. Latin American
Antiquity, 20 (2), 333-342.
Varela, H. H., y Cocilovo, J.A., 2011,. Divergencia fenotípica en los
Oasis de San Pedro de Atacama, Norte de Chile. Estudios Atacameños, 4, 101-112.
Varela, H. Cocilovo, J.A., Baffi, I. y Valdano, S., 1999, La población
antigua de la Quebrada de Humahuaca y sus relaciones biológicas con áreas
aledañas. Rev. Esp. Antrop. Biol. 20,7-24
Varela, H., Cocilovo, J.A., y Costa, M. A. 1990. El dimorfismo sexual en
la población prehistórica de San Pedro de Atacama, Chile. Chungara, 24/25:
159-166.
Varela, H.
H., Cocilovo, J.A. and Fuchs, M.L. 2017 Dispersion, Colonization and Phenotypic
Structure of Prehistoric Human Populations of the Andes of north-western
Argentina. Manuscrito.
Varela, H.H.,
Cocilovo, J.A., Fuchs, M.L. and O’Brien, T.G., 2014, Coastal Valley and Oasis
Interaction: Impact on the Evolution of Ancient Populations in the South
Central Andes. American Journal of Physical
Anthropology 155,591–599.
Varela, H.H., Cocilovo, J.A., y O´Brien, T.G., 2012, Evolución de las
Poblaciones Prehistóricas del Área Andina Centro Sur: Deriva Genética,
Migración y Selección Natural. Antropo, 28, 9-22. www.didac.ehu.es/antropo.
Varela, H.
H., Cocilovo, J. A., O’Brien, T. G. 2013. Valley to valley: the biological
connection between prehispanic residents of Cochabamba, Bolivia, and Azapa,
Chile. Advances in Anthropology, 3(04), 210.
Varela, H.H.,
O’Brien, T.G., Cocilovo, J.A., 2008, The genetic divergence of prehistoric
populations of the South-Central Andes as established by means of craniometric
traits. American Journal of Physical Anthropology 137, 274-282.
Wilder, H.,
1920, Laboratory Mannual of Anthropometry. Editorial P. Blakiston's Son y Co.,
Philadelphia.
Yacobaccio, H.D., 2012, Intercambio y Caravanas de Llamas en el Sur
Andino 3000-1000 AP. Comechingonia. Revista de Arqueología 16,31-51.