
Bernis, C., Varea, C., Terán, J.M., 2016. Ciclo
vital, transformación ambiental y las estrategias de la Historia de vida.
Antropo, 36, 29-38. www.didac.ehu.es/antropo
Ciclo vital, transformación ambiental y
las estrategias de la Historia de vida
Life
cycle, environmental transformation and Life history strategies
C.
Bernis, C. Varea y J. M. Terán
Comisión Docente de Antropología Física,
Departamento de Biología, Facultad de Ciencias, Edificio de Biológicas, Calle
Darwin 2, B-110, Universidad Autónoma de Madrid 28049 Madrid
Autora encargada de la comunicación: C. Bernis. Comisión Docente de Antropología Física,
Departamento de Biología, Facultad de Ciencias, Edificio de Biológicas, Calle
Darwin 2, B-110, Universidad Autónoma de Madrid 28049 Madrid. e-mail: cristina.bernis@uam.es
Palabras clave: Plasticidad,
Epigenética, Estrategias reproductoras, Proceso biocultural
Keywords: Plasticity, Epigenesis, Reproductive strategies, Biocultural processes.
Resumen
El Objetivo del presente trabajo es revisar la expresión
fenotípica de los procesos biológicos que ocurren a lo largo del ciclo vital asociadas
a la reciente transformación ambiental ligada a la globalización. Para ello se
revisan y discuten las evidencias que ligan transformación ambiental con
cambios en la expresión fenotípica de procesos biológicos implicados en la
reproducción. Se discuten causas y posibles consecuencias. Se detectan cambios
en la expresión de los procesos biológicos con consecuencias morfológicas, fisiológicas
y funcionales en todas las etapas de la vida, cuya duración absoluta y relativa
también se ve afectada. Se discuten su interrelación, sus tendencias de futuro
y sus consecuencias. La mayoría de los ajustes al acelerado cambio ambiental se
hacen a través de respuestas plásticas, que son continuas y se expresan dentro
de los límites biológicos impuestos por nuestros genes. La gran ventaja
adaptativa está en la maximización de la plasticidad en humanos, no necesariamente
entre los diferentes fenotipos resultantes, que varían de forma ondulante, como
el ambiente. El cambio ambiental ha sido mucho mayor en las mujeres, a través
de las mejoras en igualdad de género, que han revolucionado los comportamientos
reproductores, reduciendo la fertilidad por debajo de los niveles de
reemplazamiento y cuyas consecuencias están por evaluar.
Abstract
The objective of this paper is to revise
the phenotypic expression of the biological processes that take place
throughout the life cycle, associated with the recent environmental change due
to globalisation. With this aim, revision of papers connecting environmental
change to changes in the phenotypic expression of biological processes involved
in reproduction. Possible causes and consequences are discussed. This
evaluation shows that changes in the expression of biological processes are
detected, with morphological, physiological and functional changes in all
stages of life, whose absolute and relative length is affected. Their
interrelation, future trends and consequences are discussed. Most adjustments
to accelerated environmental change are made by plastic responses, which are
continuous and expressed within the biological limits imposed by our genes. The
great adaptive advantage is the maximisation of plasticity in humans, not
necessarily among the different resulting phenotypes, which change in a wave
pattern, like the environment. Environmental change has been much greater in
women, due to improvements in gender equality, which have revolutionised
reproductive behaviour, reducing fertility to below replacement levels, the
consequences of which are yet to be evaluated.
(…)
Let us remember that although our minds have raced ahead, our bodies have remained
much as they always were. (Short, 1994)
Introducción
El
ciclo vital está genéticamente programado y ambientalmente limitado, es el
marco de referencia natural que define y permite comprender la biología de una
especie, y se define por su duración potencial, por el número, extensión y
características de sus etapas de desarrollo y envejecimiento, por la expresión
fenotípica sus características anatómicas, fisiológicas, metabólicas y
conductuales, y por la singularidad de los patrones reproductores. La
reproducción es el mecanismo biológico que asegura la perpetuación de las
especies, de manera que todas las estructuras y funciones corporales y patrones
de comportamiento están imbricados para optimizar su reproducción en ambientes
dados.
Nuestro ciclo vital se caracteriza por su longevidad, por el
nacimiento de crías muy inmaduras que se destetan relativamente temprano, y por
un prolongado periodo de crecimiento y desarrollo distribuido en seis etapas (Bogin
y Smith, 1996), dos prenatales (embrionaria y fetal) y cuatro postnatales
(infancia, niñez, juventud y adolescencia), a lo largo de las cuales se
expresan de manera diferencial los fenotipos dependiendo de los condicionantes
ambientales (Ellison, 2001). La maduración sexual es tardía y, en las mujeres,
la vida reproductiva limitada, porque su fecundabilidad empieza a disminuir una
década antes de la menopausia sin señal de envejecimiento fisiológico y pierden
la capacidad reproductora cuando les queda por delante un tercio de su vida
potencial. El tamaño del cerebro y el tamaño y la composición corporales están
ligados al metabolismo basal y son determinantes esenciales de la Historia de
vida de los organismos (Varea y Bernis, 2013) dado que se configuran a lo largo
del desarrollo y determinan el monto total de recursos metabólicos necesarios y
su distribución (Charnov, 1993; Kuzawa, 2007).
La variabilidad fenotípica del ciclo vital humano es el
resultado de la interacción entre dos tipos de respuestas, biológicas y
culturales, que permiten el ajuste de individuos y/o poblaciones a las
condiciones ambientales que caracterizan el ecosistema que ocupan. Las
respuestas biológicas varían en función de los mecanismos implicados, de la
escala de tiempo en la que actúan y de si lo hacen sobre individuos o poblaciones.
La selección natural es la respuesta
genética frente a situaciones ambientales concretas, que moldea la
estructura genética de las poblaciones a través de mortalidad y fertilidad
diferenciales, actuando durante prolongados
periodos de tiempo y conjuntamente con el resto de mecanismos implicados
en la transmisión diferencial de genes, incluida la migración, tal y como establece
la teoría sintética de la evolución,
La plasticidad es la capacidad de los individuos de ser
moldeados por el ambiente ajustando la expresión de sus procesos biológicos o
sus comportamientos a los ambientes cambiantes en los que desarrollan sus
ciclos vitales (Lasker, 1969). La plasticidad es una propiedad compartida por
mamíferos y otros seres vivos que se maximiza en humanos (Pigliucci, 2001), que
está regulada por factores neurohormonales capaces de actuar sobre el
epigenoma, alterando los patrones de metilación del ADN, de manera que
silencian o activan la expresión génica sin modificar su estructura (Kuzawa,
2012; Jasienska, 2013). Estos cambios epigenéticos proporcionan un gran
dinamismo en la expresión génica plasmada en los fenotipos (incluyendo
información ambiental en la expresión de los genotipos), de manera que algunos
patrones de metilación epigenética se pueden mantener a lo largo de
generaciones (Drake et al., 2004), mientras
que otros se pueden modificar a lo largo del ciclo de vida de los individuos
(Ge et al., 2015).
Lo que ocurre en cada etapa de la vida, especialmente en las
tempranas, determina lo que va a ocurrir desde el punto de vista biológico y de
salud en las siguientes, de manera que las etapas de desarrollo funcionan como
un bioensayo durante el cual los
organismos reciben información ambiental y, a través de respuestas biológicas y
o culturales, ajustan, además de sus comportamientos, su tamaño y composición
corporales y la funcionalidad de órganos y sistemas (incluido el reproductor) a
las condiciones del medio en las que muy probablemente se reproducirán (Gluckman
et al., 2007; Ellison, 2001). La
capacidad de respuestas plásticas es mayor en las etapas tempranas, caracterizadas
por una rápida velocidad de crecimiento, de tal manera que se ha sugerido que
también durante la adolescencia podrían ocurrir respuestas plásticas, al menos
para el sistema reproductor, cuya máxima velocidad de crecimiento y maduración
ocurre en esa etapa (Ellison, 2001).
Las
respuestas culturales incluyen cambios en comportamientos que pueden conferir ventajas
en determinadas situaciones ambientales (por ejemplo, reducir el juego en la
infancia o el esfuerzo físico en los adultos en situaciones de privación
energética) o incluso cuestionar los modelos teóricos de la evolución, porque
se está invirtiendo la relación entre éxito reproductor y estatus (Huber et al., 2010), reduciéndose la
fertilidad por debajo de los niveles de reemplazamiento. Estas tendencias se
han interpretado como respuestas adaptativas al nuevo ambiente creado por el desarrollo
económico (Newson, 2009), aunque nosotros lo asociamos con el progreso en
igualdad social y de género, así como en la disponibilidad de métodos seguros y
eficaces de control de la fertilidad. La trasformación de las respuestas
culturales implica la transmisión generacional y/o transversal del conjunto de valores,
conocimientos, ideas, objetos y acciones que singularizan a grupos y sociedades,
actuando tanto en individuos como en poblaciones.
Material y Métodos
Se revisan trabajos propios y de otros autores que ligan
transformación ambiental con cambios en la expresión fenotípica de procesos
biológicos implicados en la reproducción y se discuten sobre causas y posibles
consecuencias.
Resultados
Historia de vida: la
energía es la clave
El principio fundamental de la Ecología evolutiva o Historia
de vida modeliza las estrategias utilizadas a lo largo de los ciclos vitales
para ajustar su variabilidad fenotípica en términos de diseños evolutivos y
optimizar la eficiencia reproductora en función de las limitaciones energéticas
que les imponen los nichos ecológicos que ocupan (Charnov, 1993). Este
principio plantea que a lo largo del ciclo vital los individuos distribuyen la energía
disponible en mantener las funciones vitales, desarrollar actividad física
cotidiana y acumular energía en forma de grasa. Una vez asegurada la
viabilidad, el resto de energía disponible se utiliza en crecer y madurar durante
las etapas de desarrollo y en reproducirse en la etapa adulta. El modelo
permite un buen ajuste de esas estrategias en todas las especies salvo en la
nuestra (Reiches et al., 2009), por
la elevada inversión energética en reproducción que deben hacer las mujeres al transformar
energía en
descendientes de grandes cerebros, que deben amamantar al menos durante
dos años de rápido crecimiento cerebral (es el órgano energéticamente más costoso
energéticamente de hacer crecer y mantener) y desarrollar su actividad física
cotidiana acarreando una criatura con más de tres kilos de peso durante gran
parte de sus vidas reproductoras.
La clave está en que la
reproducción humana es un proceso biocultural, diseñado para que otros miembros
del grupo contribuyan al gasto energético de la reproducción a través de
diferentes comportamientos (aporte de nutrientes, cuidado de lactantes y niños,
sustitución de la madre en determinadas actividades físicas cotidianas). Se han
propuesto diferentes modelos con importantes matices sobre los cuidados
alopatentales en nuestra especie y sobre quiénes son los miembros que más
contribuyen: ¿abuelas, padres, hijos mayores, población no reproductora? (Reiches et al., 2009; Coxworth et al., 2015).
Cambio ambiental reciente
Las características específicas del ciclo vital humano
construidas sobre las de nuestro pasado mamífero, primate y hominino (Ellison,
2001, 2003) quedaron establecidas hace al menos unos 200.000 años con la
aparición de la especie Homo sapiens anatómicamente
moderno, la cual, durante 8.000 generaciones, se adaptó a un ecosistema de caza
y recolección que moldeó el complejo genotípico que todavía conservamos
prácticamente invariable (Rosenberg y Trevathan, 2002; Gluckman et al., 2007; Jasienska, 2013), a pesar
de las tres grandes revoluciones ambientales ocurridas desde entonces: las que
determinaron la aparición de los ecosistemas agrícolas (hace 12000-14.000 años
y 560 generaciones), de los ecosistemas industriales-urbanos (hace 165 años y
menos de siete generaciones) y de los recientes ecosistemas urbanos virtuales del
mundo globalizado, ligada a las nuevas tecnologías e iniciada hace menos de 25
años. La transformación ambiental y de los estilos de vida resultantes se
acelera en cada uno de esos momentos y, a partir de la extensión de los
ecosistemas urbanos-industriales, se extienden tres circunstancias completamente nuevas en nuestra historia
biológica, cuyos efectos se suman en la expresión de la biología de nuestro
ciclo vital:
1. La elevada y constante disponibilidad energética y nutricional por
aumento de ingesta y gran reducción del gasto. La actividad física elevada y
cotidiana formaba parte de la ecología nutricional que singularizaba el estilo
de vida de cazadores-recolectores y, probablemente, también de agricultores
actuales no mecanizados (Malina y Little, 2008). Situaciones de elevada
actividad física disminuyen la concentración de estrógenos ováricos,
aumentándola cuando se reduce (Jasienska, 2013).
2. El progreso del conocimiento científico y técnico, y su extensión a la
sanidad pública universal, que redujo extraordinariamente la mortalidad
materno-infantil y, más recientemente, ha contribuido a la medicalización de
los procesos biológicos, especialmente los relacionados con la reproducción,
como el parto y la menopausia.
3.
Los avances en
igualdad de clase y género, que aproximan los condicionantes ambientales,
comportamientos y capacidad de decisión entre poblaciones, grupos sociales y sexos,
con importantes consecuencias sobre la fertilidad y la familia.
Estos cambios ambientales y sus
consecuencias se modelizaron por separado en las llamadas Tres transiciones, estrechamente relacionadas entre sí, pero planteadas
desde áreas de conocimiento muy diferentes: Demográfica,
centrada en los cambios en fertilidad, mortalidad y crecimiento poblacional; Epidemiológica, centrada en los cambios
de las causas y edades de muerte; y Nutricional, centrada en los cambios de disponibilidad energética y
nutricional que afectan a los procesos de desarrollo y maduración, aumentando
el riesgo de sobrepeso, obesidad y enfermedades metabólicas asociadas (Popkin,
2004) (Figura 1).
Los procesos de cambio señalados, acelerados en el
último tercio del siglo XX, no presentan una estricta coincidencia en su
desarrollo temporal ni entre poblaciones ni para una misma población. Sin
embargo, de manera esquemática se puede considerar un momento pre-transicional,
uno transicional y uno post-transicional. En las poblaciones en estadios
transicionales, se acumulan problemas socioeconómicos y biosanitarios de las
situaciones precedentes con los derivados del cambio ambiental reciente,
conviviendo problemas de sobrepeso, enfermedades metabólicas y cardiovasculares
con elevada carga infecciosa y de desnutrición, elevadas pérdidas infantiles y
deficiente estado biológico y de salud (Custodio et al., 2009). Debemos ser conscientes de que la transformación
ambiental es continua y ondulante, con tendencias direccionales durante
prolongados periodos de tiempo, y de que nunca retornaremos a los estilos de
vida de nuestros antecesores cazadores-recolectores, con una limitada
disponibilidad energética, una elevada actividad física cotidiana y una
fertilidad elevada distribuida a lo largo de un periodo reproductor potencial
más reducido que el nuestro. Pero podemos beneficiarnos de sus experiencias si
comprendemos por qué nuestro sistema reproductor evolucionó como lo hizo
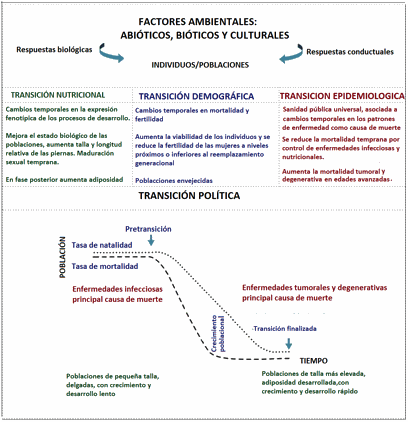
Figura 1. Transformación ambiental reciente (1850-1990)
Figure 1. Recent enviromental change (1850-1990)
Consecuencias del cambio
ambiental sobre los procesos reproductores
Los determinantes biológicos de la reproducción humana, incluidas
las diferencias sexuales, se han establecido a lo largo de la evolución para
optimizar el esfuerzo reproductor. Los determinantes culturales —que
presentan una gran diversidad poblacional y un acelerado cambio temporal (Bernis,
1999; Varea, 2009)— interactúan con los biológicos modulando su maduración y desarrollo, la
utilización de su potencial reproductor, y las decisiones sobre nutrición,
protección y cuidados a los descendientes. Los principales cambios en la
expresión de la biosociología reproductiva afectan a tres aspectos
esenciales: a) los relacionados con el parto, que limita la etapas pre y post-natal;
b) los asociados con las edades de menarquia y menopausia, que limitan las
etapas pre y post- reproductora de la reproductora; y c) los relacionados con
la fertilidad, que mide la eficiencia biológica de los individuos y la
capacidad de crecimiento poblacional.
Los
determinantes culturales han sido y siguen siendo los principales moduladores
del acceso a energía y nutrientes que influyen sobre los procesos fisiológicos
de la reproducción y sobre el número final de descendientes. El acceso
diferencial de energía y nutrientes se asocia con la limitación de recursos por
género y nivel socioeconómico (menos ingesta y mayor exigencia de esfuerzo
físico desde la infancia y durante el embarazo en los más desfavorecidos).
Además, los comportamientos culturales
definen los patrones reproductores en cada población, regulando su participación
o exclusión del juego reproductor, las decisiones sobre la elección de la
pareja, sobre su número y sus características (por ejemplo, parentesco, edad de
incorporación y de finalización del proceso reproductor), sobre el número ideal (socialmente adecuado) de
descendientes, y sobre las decisiones y capacidades técnicas para mantenerlos
vivos y proteger su salud hasta edad reproductora.
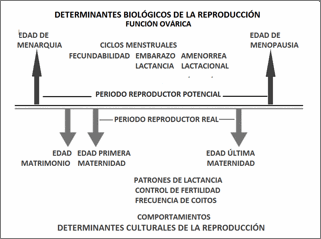
Figura 2. Determinantes
biológicos y culturales de la reproducción humana
Figure 2. Biological and
cultural determinants of Human reproduction.
Los determinantes biológicos regulan
los momentos de inicio y finalización del periodo fértil de las mujeres a
través de las edades de menarquía y de menopausia, estableciendo, por un lado,
la duración absoluta del periodo reproductor potencial y, por otro, las
características fisiológicas y funcionales de los ciclos menstruales, del útero
y de los ovarios, capaces estas de ajustarse de manera puntual o prolongada a
situaciones de estrés energético a lo largo de la vida reproductiva, como
demostró Ellison (2003), padre de la Ecología reproductiva. Se genera así
variabilidad en la probabilidad de concebir, de llevar a cabo un embarazo, del
número de embarazos potenciales y del número de recién nacidos vivos. La
funcionalidad y la tardía diferenciación de las glándulas mamarias solo se
completan tras la primera lactancia, es decir depende de la edad de primera
maternidad que tiene un fuerte control cultural. Finalmente, la expresión de
los determinantes biológicos a lo largo del desarrollo determinan el estado
biológico en la etapa reproductora, que modula la bondad del desarrollo fetal y
del recién nacido, y la posterior inversión parental necesaria para proteger a
los descendientes (Figura 2).
Las consecuencias del cambio
ambiental reciente sobre los determinanates biológicos de la reproducción se
pueden resumir en la tendencia hacia menarquias más tempranas y menopausias más
tardías, lo que aumenta el periodo reproductor potencial. La edad de la primera primera
regla marca las caracteristicas de la función ovárica, de tal manera que la
maduración temprana se asocia con la temprana regularización de los ciclos ovulatorios y elevadas
concentraciones de estrógenos durante la vida fértil (Apter et al., 1978; Ellison, 2001; Vitzthum,
2009; Jasienska, 2013). Por su parte, la
edad de menopausia y las características de la perimenopausia también están
ligadas a las circunstancias ambientales que modulan el aporte energético en etapas
previas (así como al tabaquismo) (Varea et
al., 2000). Poblaciones con peores circunstancias socioeconómicas,
nutricionales y un exigente trabajo físico tienen menopausias más tempranas con
peri-menopausias más cortas y menos sintomáticas. La edad de menopausia es más
tardía en mujeres con una reducida o nula actividad física, elevadas ingestas
energéticas y un IMC en el rango de la obesidad, tendiendo además transiciones
más largas, que refieren mayor frecuencia y número de síntomas diferentes (no
solamente vasomotores) y recurren más a terapia hormonal sustitutiva y a
soluciones quirúrgicas (Bernis y Reher, 2007). El resultado es un importante
aumento del número de ciclos menstruales ovulatorios que Eaton et al. (1994) estiman en unos 450 ciclos
por término medio en la mujer estadounidense, frente a 160 en mujeres cazadoras-recolectoras
contemporáneas. Mujeres rurales y urbanas de Marruecos estudiadas entre 1980 y
1990 con elevados tamaños familiares (7,2 y 8,5 nacidos vivos) y lactancias
prolongadas tenían unos 150 ciclos (Varea et
al., 1996). Mujeres rurales españolas, con un tamaño familiar medio de 3,3 nacidos
vivos y lactancias prolongadas en la década de los años 70, experimentaban unos
286 ciclos, mientras que mujeres urbanas estudiadas en los años 90, con tamaños
familiares medios de 2,8 nacidos vivos y lactancias muy reducidas, el número de
ciclos aumenta a 411 (Bernis, 1999). El elevado número de ciclos menstruales
con elevados niveles de estrógenos que hoy día tienen las mujeres aumenta el
riesgo de padecer cáncer de mama, especialmente si tenemos en cuenta la
importante reducción temporal de la actividad física que reduce los niveles de
estrógenos y es un elemento protector (Eaton et al., 1994; Jasienska, 2013).
Estado biológico en las etapas
tempranas: desarrollo fetal, parto y recién nacido
El parto es el momento transicional más
corto y más peligroso del ciclo vital humano, que separa las dos etapas con mayor
diferencia entre el ambiente previo y posterior. La concepción inicia el
complejo reajuste del metabolismo energético que caracteriza cada suceso
reproductor, siendo el embarazo el periodo energética y nutricionalmente más sensible del ciclo vital, el que presenta
la máxima
plasticidad metabólica y cuyo desarrollo puede tener más consecuencias a largo
plazo sobre el estado biológico y de salud en la etapa reproductora y en las
edades más avanzadas. El feto puede ajustar permanentemente
un metabolismo más eficaz a situaciones de malnutrición materna porque así se prepara para una vida postnatal que
supuestamente tendrá las condiciones energéticas próximas a las experimentadas
en el ambiente materno. Muchas poblaciones de los siglos XX y XXI —incluida
la española— han estado sometidas a una importante transformación ambiental
entre generaciones sucesivas, de manera que se gestaron y fueron niños en
circunstancias de deprivación energética, si bien luego desarrollaron sus vidas en las circunstancias
opuestas ya comentadas, lo que aumentó su riesgo de desarrollar determinadas
enfermedades, fisiológicas y metabólicas en adultos y
ancianos (Barker,
1994;
Ellison, 2001). Hoy sabemos que el peor escenario para las perspectivas
individuales de salud metabólica y fisiológica se asocia precisamente con
desajustes importantes entre los ambientes fetal y adulto, mientras que la
coincidencia entre ambos, sea rico o pobre en ambas etapas, reduce el riesgo de
esas enfermedades (Jasienska, 2013).
Nacer en condiciones biológicas óptimas es
un pasaporte de salud para el resto de la vida, y está muy condicionado por las
características que definen al denominado Sistema materno (WHO, 2012). A pesar del indiscutible progreso en el estado biologico de
las madres y de los recién nacidos detectados hasta 1980 aproximadamente y de los
considerables progresos en investigación y salud pública, todavia nacen anualmente
15 million de bebés pretérmino (antes de la semans 37 de gestación), un millón de
los cuales muere de complicaciones derivadas de ello, mientras que muchos de
los que se mantienen vivos sufren discapacidad permanente. Lo más preocupante
es que desde finales del siglo XX, hay una inversión en las tendencias de los
nacidos prematérmibo y con bajo peso al nacer (BPN, menos de 2.500
gramos), que aumentan en la mayoría de los
paises desarrollados y en desarrollo (WHO, 2012; EURO-PERISTAT Project, 2013), generando además un
importante coste sanitario. En madres españolas se ha detectado el mismo
fenómeo desde 1980, de manera que, independientmente de las diferencias entre
el peso de nacidos vivos, nacidos muertos y muertos menores de un día, en todos
ellos ha disminuido su peso medio al nacer tanto en partos únicos como en
múltiples (Bernis, 2010) sin que haya aumentado los partos prematérmino, como
sí ha ocurrido en otros países europeos (EURO-PERISTAT
Project, 2013). Ello sugiere que algo no está funcionando en la prevención
de salud perinatal (Bernis et al., 2012; Varea et al., 2014). El análisis de las diferencias en el estado
biologico de los recien nacidos y de sus tendencias entre 1996 y 2010 en
función del origen materno (con una contribución máxima de madres extranjeras
en 2008 del 21%) porporciona una información complementaria que abre nuevos
interrogantes. A lo largo del periodo analizado, el peso medio y el peso por
edad de gestación de los los recién nacidos de madre extranjera aumentó a lo
largo del periodo considerado y fué significativamente mayor que el de los de nacidos
de madres españolas. Entre los hijos de españolas y hasta el nicio de la crisis
económica iniciada en 2008, aumentaron los recién nacidos con BPN (tanto en
partos a término como en pretérmino) y disminuyó el peso medio al nacer (tanto de
partos multiples como únicos), mientras que el peso por edad de gestación no
experimentó cambio temporal. Ambos grupos materno-fetales comparten una
reducción de la edad de gestación, menos acentuada en las extranjeras, que
determina un aumento los embarazos a término de 37-38 semanas, lo que sugiere
una posible causa común relacionada con el el aumento de intervenciones
obstétricas (Bernis et al., 2012), como se ha
sugerido también para otras poblaciones europeas (EURO-PERISTAT Project, 2013). A ello se suma, primeras evidencias de un impacto negativo
de la actual crisis económica sobre el crecimiento fetal en nacidos de madres
españolas (Varea et al., 2016).
Conclusiones
La cultura es el principal agente de
cambio ambiental que ha revolucionado los estilos de vida, estilos que afectan
directa o indirectamente a la expresión fenotípica de los procesos biológicos,
especialmente los relacionados con la disponibilidad energética, con el
conocimiento científico y su aplicación a través de una sanidad pública, y con
la igualdad de clase y género. Esta nueva situación en nuestra historia
biológica se puede resumir diciendo que nos sitúa en una dirección contraria a
los modelos teóricos clásicos, al combinar una permanente y excesiva
disponibilidad energética (que modifica nuestra edad de maduración sexual y
nuestra fisiología ovárica, nuestro desarrollo fetal y el estado biológico del
recién nacido, y el riesgo de enfermedad tumoral) con un cambio radical en las decisiones sobre utilización de la
vida reproductiva (fertilidad nula o baja, maternidad tardía). Es decir, la interacción entre
respuestas fisiológicos y conductuales afecta al éxito reproductor (porque se
está reduciendo la fertilidad por debajo de los niveles de reemplazamiento) y a
la salud de las poblaciones (porque aumentan sobrepeso y obesidad en todas las
edades y el riesgo de padecer determinadas enfermedades metabólicas y
cardiovasculares tanto por exceso de disponibilidad energética a lo largo de la
vida, como por el aumento del desajustes entre el ambiente fetal y el adulto: Jasienska,
2013).
En cualquier caso, nuestra vida reproductiva nunca volverá a
ser como la de nuestros antepasados cazadores-recolectores y desde las ciencias
biomédicas se buscan soluciones para los problemas expuestos, resultantes de
las limitaciones de nuestros genes —que mantienen un cuerpo anatómica y
funcionalmente igual al de los primeros miembros de nuestra especie— y
las exigencias del ambiente cultural en rápido cambio. ¿Podemos, entonces,
predecir nuevas tendencias en la biología del ciclo vital derivadas de la rápida
y continua transformación ambiental, y evaluar sus consecuencias a medio plazo
sobre la salud y tomar decisiones racionales sobre la manera de ajustarnos para
mejor a las nuevas demandas que nuestro ambiente cultural nos está imponiendo?
La respuesta es compleja y requiere que la perspectiva evolutiva se incorpore a
la investigación biomédica, y deja numerosas cuestiones abiertas (Haig, 2008;
Kuzawa, 2012). La evolución no actúa seleccionando individuos globalmente sanos
porque el cuerpo y su funcionamiento a lo largo de la vida son el resultado de
un conjunto de soluciones de compromiso. Y esto es así porque la mayoría de los
ajustes al acelerado cambio ambiental se hace a través de respuestas plásticas que
son continuas y se expresan dentro de los límites biológicos impuestos por
nuestros genes, por encima y por debajo de los cuales la viabilidad no es
posible, produciéndose primero patologías o disfunciones y, finalmente, si no
mejoran las circunstancias, la muerte. En cualquier caso, la gran ventaja
adaptativa está en la maximización de la plasticidad en humanos, no
necesariamente entre los diferentes fenotipos resultantes de la misma. La pregunta
—todavía sin respuesta— es si en las nuevas circunstancias de
rapidísimo cambio ambiental y conductual se están sobrepasando los límites de
nuestra plasticidad impuestos en los ambientes en que fuimos seleccionados.
Bibliografía
Apter,
D., Viinikka, L., Vihko, R., 1978. Hormonal pattern of adolescent menstrual cycles. Journal of Clinical
Endocrinology and Metabolism, 47: 944-954.
Barker, D., 1994. Fetal origins of coronary heart disease. British
Medical Journal, 311: 171-174.
Bernis, C., 1999. Changing lifestyles and reproductive health. En Health
Ecology. Health, culture and human-environment interaction, editado por M. Horani,
y T. Boleyn, (Londres y Nueva York: Routledge) p. 153.
Bernis,
C., Reher, D.S., 2007. Environmental contexts of menopause in Spain:
comparative results from recent research. Menopause, 14: 1-11.
Bernis, C., 2010. Factores causales de la
reducción del peso al nacer en España 1980-2007: cambios en la viabilidad
fetal, en la distribución de la edad gestacional y en la dinámica del
crecimiento fetal. Trabajos de Antropología, 31: 233-247.
Bernis, C., Varea, C., Bogin, B.,
González-González, A., 2012. Labor management and
birth outcome among migrant and Spanish women in Madrid: the effect of changing patterns of obstetric care on rates of preterm
and low birth weight. Maternal
and Child Health Journal, 17: 918-927.
Bogin, B., Smith, B.H., 1996. Evolution of the human life cycle. American
Journal of Human Biology 8, 703-716.
Charnov, E.L., 1993. Life history invariants: some explorations symmetry
in the evolutionary ecology. (Oxford: Oxford University Press). pp. 167.
Coxworth,
J.E., Kim, P.S., McQueen, J.S., Hawkes, K., 2015. Grandmothering life histories
and human pair bonding. Proceedings of the National Academy of Sciences of the
United States of America, 112: 11806-11811.
Custodio, E., Descalzo, M.Á., Roche, J.,
Molina, L., Sánchez, I., Lwanga, M., Torres, A. M., Fernández-Zincke, E., Bernis,
C., Villamor, E., Baylin, A., 2010. The economic and
nutrition transition in Equatorial Guinea coincided with a double burden of
over- and under nutrition. Economics and Human Biology, 8: 80-87.
Drake, A.J., Walker, B.R., 2004. The
intergenerational effects of foetal programming: non-genomic mechanisms for the
inheritance of low birth weight and cardiovascular risk. Journal of Endocrinology, 180: 1-16
Eaton,
S.B., Pike, M.C., Short, R.V., Lee, N.C., Trussell, J., Hatcher, R.A., Wood,
J.W., Worthman, C.M., Jones, N.G.B., Konner, M.J., Hill, K.R., Bailey, R.,
Hurtado, A. M., 1994. Women's reproductive cancers in evolutionary context.
Quarterly Review of Biology, 69: 353-367.
Ellison,
P.T., 2001. On Fertile Ground. A Natural History of Human Reproduction.
(Cambridge, MA: Harvard University Press). pp. 346.
Ellison,
P.T., 2003. Energetic and reproductive effort. American Journal of Human
Biology, 15:342-351.
EURO-PERISTAT Project. 2013. European Perinatal Health Report. The
health and care of pregnant women and babies in Europe in 2010. Paris: Euro-Peristat.
pp 252.
Ge, Z.J., Schatten, H., Zhang, C.-L., Sun, Q.-Y., 2015. Oocyte ageing
and epigenetics. Reproduction 149: R103-R114.
Gluckman, P.D., Hanson, M.A., Beedle, A.S., 2007. Early life events and
their consequences for later disease: a life history and evolutionary
perspective. American Journal of Human Biology, 19: 1-19.
Haig, D., 2008. Intimate relations: Evolutionary conflicts of pregnancy
and childhood. Evolution in Health and disease, editado por S.C. Stearns y J.C.
Koella (New York: Oxfors University Press) p. 65.
Huber, S., Bookstein, F.L., Fieder, M., 2010. Socioeconomic Status,
Education, and Reproduction in Modern Women: An Evolutionary Perspective. American Journal of Human Biology. 22:
578-587.
Jasienska, G., 2013. The fragile Wisdom. An evolutionary view on women´s
biology and health (Harvard University Press). pp 336.
Kuzawa, C., 2007. Developmental origins of life history: growth
productivity and reproduction. American Journal of
Human Biology, 19: 654-661.
Kuzawa, C., 2012. Why evolution needs development, and medicine needs
evolution. International Journal of Epidemiology, 41: 223-229.
Lasker, G.W., 1969. Human biological adaptability. Science,
166: 1480-1486.
Malina, R.M., Little, B.B. 2008. Physical Activity: The Present in the Context of the Past. American Journal
of Human Biology, 20: 373-391.
Newson, L., 2009. Cultural Versus Reproductive Success: Why Does
Economic Development Bring New Tradeoffs? American Journal of Human Biology, 21:
464-471.
Pigliucci, M., 2001. Phenotypic plasticity: beyond nature and nurture (Baltimore:
John Hopkins University press). pp 328.
Popkin, B.M., 2004. The nutrition transition an overview of world
patterns of change. Nutrition Reviews, 62: S140-S143.
Reiches, M.W., Ellison,
P.T., Lipson, S.F., Sharrock, K.C., Gardiner, E., Duncan, L.G., 2009. Pooled energy budget and human life history. American Journal of Human
Biology, 21: 421-429.
Rosemberg, K. Trevathan, W., 2002. Birth, obstetrics and human
evolution. British Journal of Obstetrics and Gynaecology, 109: 1199-1206.
Short, R,V., 1994. Human reproduction in an evolutionary context. En
Human Reproductive Ecology: Interactions of Environment, Fertility and
Behavior, editado por Kenneth L Campbell, y James W Wood (Nueva York: New York
Academy of Sciences) pp. 416-425
Varea, C., Bernis,
C., Montero, P., Arias, S., Barroso, A., González, B. 2000. Secular trend and intrapopulational variation in age at menopause in
Spanish women. Journal of Biosocial Science, 32: 383-393.
Varea, C., Crognier, E., Bley, D., Boëtsch, G., Baudot, P., Baali, A.,
Hilali, M.K., 1996. The determinants of contraceptive use in Morocco: stopping
behavior in traditional populations. Journal of Biosocial Science, 28: 1-13.
Varea, C.. 2009.
El debate sobre un nuevo patrón reproductor en España y la contribución del colectivo
de mujeres inmigrantes. La maternidad en el siglo XXI. Mitos y realidades,
editado por C. Bernis, R.López y P. Montero (Madrid: Ediciones de la
Universidad Autónoma) p. 171.
Varea, C., Bernis, C.. 2013. Encephalization, Reproduction and Life History.
Human Evolution 28: 1-16.
Varea, C., Bernis, C., González-González, A., 2014. Biosocial aspects of
fetal growth and birth outcome. Human Growth: The Mirror of the Society,
editado por M. Sidkhar (Delhi: BR Publishing Corporation) p. 196.
Varea, C.,
Terán, J.M., Bernis, C., Bogin, B., González-González, A., 2016. Is the economic crisis affecting birth outcome in Spain? Evaluation of
temporal trend in underweight at birth (1996-2012). Annals of Human Biology, 43:
169-182.
Vitzthum, V.J., 2009. The ecology and evolutionary endocrinology of
reproduction in the human female. American Journal of Physical Anthropology, 140:
S95-S136.
WHO. 2012. Born too soon: the global action report on preterm birth. (Ginebra:
WHO, Suiza). pp. 126.