Moussouni, A., Aouar Metri, A., 2011,
Etude du polymorphisme des dermatoglyphes et des groupes sanguins (ABO, Rhésus,
MNSs, Duffy et Kell) chez la population de Sabra dans le Nord Ouest Algérien. Antropo,
25, 65-80. www.didac.ehu.es/antropo
Etude du
polymorphisme des dermatoglyphes et des groupes sanguins (ABO, Rhésus, MNSs,
Duffy et Kell) chez la population de Sabra dans le Nord Ouest Algérien
Study of
polymorphism of dermatoglyphics and blood groups (ABO, Rhesus, MNSs, Duffy and
Kell) in the Sabra population in northwestern Algeria
Abdellatif Moussouni1
et Ammaria Aouar Metri2
1
Laboratoire
d’Anthropologie des Religions et comparaison, Faculté des Sciences Humaines et
Sociales, Université Abou Bekr Belkaïd de Tlemcen, Algérie. biomorice@yahoo.fr
2 Laboratoire de valorisation de l’action de l’homme pour la protection
de l’environnement et application en santé publique (équipe environnement et
santé), faculté des sciences, Université Abou Bekr Belkaïd de Tlemcen, Algérie. aaouar@netcourrier.com.
Mots clés: population, Sabra, Algérie, marqueurs sanguins, diversité génétique, polymorphisme, dermatoglyphes.
Keywords: population, Sabra, Algeria, blood markers,
genetic diversity, polymorphism, dermatoglyphics.
Résumé
La
présente étude s’intéresse à la caractérisation anthropogénétique de la
population de Sabra située à l’extrême Ouest Algérien à travers une analyse
comparative des polymorphismes des dermatoglyphes et des groupes sanguins (ABO,
Rhésus, MNSs, Duffy et Kell) avec le reste des populations Algériennes.
L’analyse des dermatoglyphes sur un échantillon de 200 personnes (100 hommes et 100 femmes) a permis de mettre en évidence une prédominance des Boucles ulnaires, suivies de près des Tourbillons, des Arcs et des Boucles radiales. L’analyse comparative des fréquences de ces figures digitales dans le contexte Méditerranéen, a montré un rattachement de notre population au groupe des populations de l’Afrique du Nord et du Moyen Orient.
L’analyse hémotypologique a été effectuée dans les systèmes ABO, Rhésus, MNSs, Duffy et Kell. Le phénotypage érythrocytaire par micro- méthodes à concerné 203 sujets pour le système ABO, 100 pour les systèmes Rhésus et Kell et 22 pour les systèmes MNSs et Duffy.
Elle a révélé les
cinq systèmes en équilibre génétique (E.H.W).
Cependant, l’analyse en composantes principales, l'estimation des distances génétiques pour l'ensemble des cinq marqueurs analysés, ainsi que les arbres phylogénétiques construits ont montré le profil génétique de notre population très proche de celui de la majeure partie des populations d’Algérie.
Abstract
This study focuses on the
characterization of population anthropogénétique Sabra located in the extreme
western Algeria through a comparative analysis of polymorphisms of
dermatoglyphics and blood groups (ABO, Rhesus, MNSs, Duffy and Kell) with the
rest of Algerian population. Analysis of dermatoglyphics in a sample of 200
persons (100 men and 100 women) has to demonstrate a predominance of ulnar loops,
followed closely eddies, arcs and radial loops. Comparative analysis of digital
frequencies of these figures in the Mediterranean context, showed an attachment
of our people to the group of populations of North Africa and the Middle East.
The analysis was performed hémotypologique systems ABO, Rhesus, MNSs, Duffy and
Kell. The micro-erythrocyte phenotyping methods involved 203 subjects for the
ABO system, 100 for Rhesus and Kell systems and 22 for MNSs and Duffy systems.
She found the five systems in genetic equilibrium (EHW). However, the principal
component analysis, estimation of genetic distances for all five markers
analyzed and the phylogenetic trees showed the genetic profile of our
population very close to that of the major part of the population of Algeria.
Introduction
L’anthropobiologie aborde tant la variabilité contemporaine que la reconstitution de leur histoire évolutive. Le terme de variabilité ou de variation est utilisé pour qualifier la différence qui existe entre les individus d’une même population ou de populations différentes. Selon la nature des caractères étudiés et de leur déterminisme génétique divers outils d’analyses sont utilisés (Charles et al., 2003).
Ainsi l’'anthropologie biologique et spécialement son versant immunogénétique s'intéresse à l’exploration du polymorphisme génétique humain modelée par les données fournies par la génétique des populations qui sont gouvernés par des paramètres qui ne sont pas tous de l’ordre génétique, puisque outre la mutation, la sélection ou la dérive génétique, ils s’intègrent les écarts à la panmixie, les migrations et la taille de la population (Loi de Hardy Weinberg). Par conséquent, l’ensemble de ces phénomènes conduit à une variation des fréquences des gènes entre les différentes populations (Terzian et Biemont, 1988).
Cette exploration permet de saisir la diversité génétique entre et à l’intérieur des différents groupes humains, compte tenu que l’Anthropologie d’aujourd’hui se préoccupe moins de classer que d’expliquer la diversité.
Au sein d’une population, la variabilité entre les individus peut provenir de leur diversité génétique et de la diversité des milieux dans lesquels s’expriment leurs gènes. Ainsi donc, l'évolution d'une population est le résultat des interactions entre son patrimoine biologique, sa culture et le comportement de ces membres.
Par ailleurs, la performance de la définition de la structure génétique d’une population repose avant tout sur l’analyse de plusieurs systèmes génétiquement indépendants les uns des autres.
D’abord, Les groupes sanguins sont des marqueurs génétiques classiques, présentant un grand degré de polymorphisme, ce qui leur donne un intérêt particulier dans les études de micro différentiation et l’histoire migratoire des peuplements (El Ossmani et al., 2008).
Cependant, avec le développement de l’anthropologie, les dermatoglyphes ne tardèrent à intéresser les chercheurs dont le souci majeur était la recherche et la détermination de la variabilité des populations humaines (Jean, 1987).
Ces figures digitales furent utilisées depuis l’antiquité comme signature en Babylone et en Chine (Pichard et al., 2004), et restent aujourd’hui encore une des méthodes d’identification les plus utilisées grâce à leur constante, leur spécificité et leur pouvoir discriminant (Cummin et Midlo, 1961).
Dans le cadre de l’étude de la variation biologique des populations actuelles, le plus recommandé est de travailler sur les populations aborigènes (Cavalli-Sforza et al., 1994).
Depuis la protohistoire, le pourtour Méditerranéen connaît un mouvement ininterrompu d’hommes et d’idées brassant ses peuples et ses cultures (Sabir et al., 2004). Cependant la population Algérienne est rattachée essentiellement aux groupes ethniques berbère et arabe, en dépit des multiples invasions qu’a connues dans le passé l’Afrique du Nord: par les Phéniciens, les Romains, les Vandales, les Arabes, les Turcs et les Français (Aireche et Benabadji, 1994).
Dans ce contexte, la population Algérienne à fait l’objet de quelques analyses hémotypologiques (Solal et al., 1952; Auzas, 1957; Ruffié et al., 1960, 1963 et 1966; Benabadji et Chemla, 1971; Aireche et Benabadji, 1988 et 1994).
Dans la même perspective et en raison de l’évolution socioculturelle de la société Algérienne, ainsi que devant la rareté des données anthropogénétiques sur les populations Algériennes nous nous sommes intéressés à combler ce vide par un nouveau travail. Ce a porté sur l’étude de la variabilité génétique et la détermination de la position anthropogénétique de la population de Sabra dans l’espace Méditerranéen, à travers l’analyse comparative de deux types de polymorphismes (dermatoglyphes digitaux et groupes sanguins ABO, Rhésus, MNSs, Duffy et Kell).
Matériel et Méthodes
L’étude a été portée sur la région de Sabra qui se situe à l’extrême Ouest d’Algérie à proximité des frontières algéro- marocaines à l’Ouest de la wilaya de Tlemcen le long de la route reliant Tlemcen 29 km et à l’Est à la ville de Maghnia 24 km Ouest. Sa superficie est estimée à 33056 hectares renfermant prés de 30700 habitants. Sur le plan découpage administratif, en plus de son chef lieu Sabra compte aussi quatre agglomérations secondaires:
Oued Ezzeitoune, Belghafer, Sidi El Arbi et Moul Eddechra ainsi que nous pouvons retrouver quelques zones dispersées formant des douars (figure 1).
L’échantillonnage
a été réalisé au sein du laboratoire d’analyses médicales de la polyclinique de
Sabra sur des individus arabophones, apparemment sains volontaires, originaires
de la région et n’ont pas de liens de parenté. Une fiche de consentement préalable a été signée
par tous les participants.
L’analyse des dermatoglyphes a été effectuée sur 200 sujets (100 hommes et 100 femmes). Pour l’obtention des empreintes digitales, nous avons utilisé la technique qui consiste à enduire la surface ridée d’encre. L’extrémité distale de chacun des dix doigts est roulée sur du papier blanc Pour chaque individu, nous avons déterminé les figures digitales situées à l’extrémité apicale des doigts de chaque main, celles-ci ont été classées en: Arcs plats (Ap), Arcs en tente (At), Boucles ulnaires (Bu), Boucles radiales (Br), Tourbillons (T) et Doubles boucles (DB). Les résultats obtenus ont été classés par doigt, main et pour la somme des deux mains aussi bien chez les hommes que chez les femmes.
Cependant, l’analyse hémotypologique a été menèe dans les systèmes ABO, Rhésus, MNSs, Duffy et Kell. Pour les prélèvements sanguins nous avons collecté à partir de chaque individu environ 10 ml de sang par ponction veineuse à l’aide d’une seringue stérile recueilli dans des tubes contenant l’EDTA comme anti- coagulant. Le phénotypage par micro- méthode a concerné 203 sujets pour le système ABO, 100 pour les systèmes Rhésus et Kell et 22 pour les systèmes MNSs et Duffy.
Analyses Statistiques
Les différences bimanuelles et sexuelles ont été évaluées à l’aide du test d’indépendance χ2 calculé par le logiciel MINITAB version 12.
Les fréquences alléliques et haplotypiques ont étédéterminées par la méthode de maximum de vraisemblance basée sur l’hypothèse d’équilibre de Hardy Weinberg. Pour vérifier ce dernier nous avons utilisé le test χ2 qui mesure l’écart entre les fréquences observées et les fréquences théoriques. Pour les comparaisons des fréquences géniques et haplotypiques de notre population avec celles des populations d’Algérie ont été effectuées par le test χ2 réalisé par le programme BIOSYS-1.
Les relations biologiques entre les populations ont été représentées également par un diagramme bidimensionnel obtenu après une analyse en composant principal (Dagnelie, 1970), est réalisé grâce au logiciel informatique ADE-4. Cependant, Les distances génétiques entre les populations ont été réalisées en utilisant les mesures standards de la variation des fréquences géniques selon le coefficient de coancestralité de Reynolds et al. (1983) avec le programme package PHYLIP 3.5 C (Felsenstein, 1989). Enfin, Les arbres phylogénétiques ont été construits à l’aide du logiciel « Neighbor Joining » (Saïtou et Neï, 1987) du programme package PHYLIP 3.6 C afin d’établir le degré de similitude entre les populations.
Pour le degré de la signification du test χ2:
P>0.05: une différence non significative (NS)
P<0.05: une différence significative (*)
P<0.01: une différence hautement significative (**)
P<0.001: une différence très hautement significative (***)
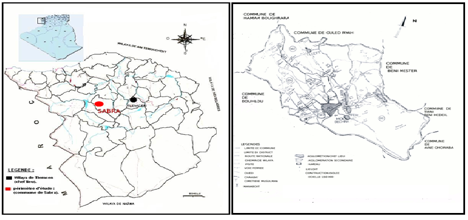
Figure 1. La carte de Tlemcen et la position de la
région de Sabra.
Figure
1.
The map of Tlemcen and the position of the Sabra area.
Résultats et discussion
Résultats des
dermatoglyphes
Les résultats obtenus concernant les fréquences des différents types de figures digitales chez la population de Sabra (tableau 1), montrent que les figures les plus fréquentes pour les deux mains chez l’ensemble de la population sont les Boucles ulnaires (51.30%) suivies de prés des Tourbillons (33.55%), des Doubles Boucles (5.80%), des Arcs plats (5%), des Arcs en tente (2.20%) et enfin des Boucles radiales (2.15%).
|
Sexe Figures |
Hommes |
Femmes |
Hommes + Femmes |
||||||
|
D |
G |
D + G |
D |
G |
D + G |
D |
G |
D + G |
|
|
Arcs plats |
5.60 |
4.20 |
4.90 |
5.00 |
5.20 |
5.10 |
5.30 |
4.70 |
5.00 |
|
Arcs en tente |
2.20 |
3.00 |
2.60 |
0.60 |
3.00 |
1.80 |
1.40 |
3.00 |
2.20 |
|
Total arcs |
7.80 |
7.20 |
7.50 |
5.60 |
8.20 |
6.90 |
6.70 |
7.70 |
7.20 |
|
Boucles ulnaires |
48.80 |
48.00 |
48.40 |
59.00 |
49.40 |
54.20 |
53.90 |
48.70 |
51.30 |
|
Boucles radiales |
2.60 |
3.40 |
3.00 |
0.60 |
2.00 |
1.30 |
1.60 |
2.70 |
2.15 |
|
Doubles boucles |
2.20 |
3.60 |
2.90 |
5.00 |
12.40 |
8.70 |
3.60 |
8.00 |
5.80 |
|
Tourbillons |
38.60 |
37.80 |
38.20 |
29.80 |
28.00 |
28.90 |
34.20 |
32.90 |
33.55 |
|
Total tourbillons |
40.80 |
41.40 |
41.10 |
34.80 |
40.40 |
37.60 |
37.80 |
40.90 |
39.35 |
Tableau 1. Fréquences des figures Dermatoglyphiques
chez la population de Sabra.
D: droit, G:gauche.
Table 1. Frequencies figures dermatoglyphics in the population
of Sabra.
D: right; G: left
Chez les deux sexes de notre population nous avons noté une fréquence relativement importante des Boucles ulnaires et des Tourbillons sur la main droite par rapport à la main gauche. A l’opposé, la fréquence des Arcs en tente, des Boucles radiales et des Doubles Boucles est principalement important sur la main gauche comparée à la main droite.
Par doigts, nous avons vu chez les hommes une prédominance des Arcs et des Boucles radiales aux doigts II, des Boucles ulnaires aux doigts V et des Tourbillons aux doigts I. Par ailleurs, chez les femmes nous avons observé également la prédominance des Arcs aux doigts
III, des Boucles radiales aux doigts II, des Boucles ulnaires aux doigts III et des Tourbillons aux doigts I (tableau 2).
|
Doigts Figures |
I |
II |
III |
IV |
V |
I - V |
||||||
|
D |
G |
D |
G |
D |
G |
D |
G |
D |
G |
D |
G |
|
|
Hommes |
||||||||||||
|
Arcs plats |
3.00 |
1.00 |
8.00 |
6.00 |
8.00 |
7.00 |
3.00 |
3.00 |
6.00 |
4.00 |
5.60 |
4.20 |
|
Arcs en tente |
0.00 |
0.00 |
5.00 |
6.00 |
4.00 |
5.00 |
1.00 |
1.00 |
1.00 |
3.00 |
2.20 |
3.00 |
|
Total arcs |
3.00 |
1.00 |
13.00 |
12.00 |
12.00 |
12.00 |
4.00 |
4.00 |
7.00 |
7.00 |
7.80 |
7.20 |
|
Boucles ulnaires |
38.00 |
43.00 |
37.00 |
36.00 |
60.00 |
56.00 |
46.00 |
51.00 |
63.00 |
54.00 |
48.80 |
48.00 |
|
Boucles radiales |
2.00 |
0.00 |
7.00 |
12.00 |
2.00 |
2.00 |
1.00 |
2.00 |
1.00 |
1.00 |
2.60 |
3.40 |
|
Doubles boucles |
10.00 |
14.00 |
1.00 |
3.00 |
0.00 |
1.00 |
0.00 |
0.00 |
0.00 |
0.00 |
2.20 |
3.60 |
|
Tourbillons |
47.00 |
42.00 |
42.00 |
37.00 |
26.00 |
29.00 |
49.00 |
43.00 |
29.00 |
38.00 |
38.60 |
37.80 |
|
Total tourbillons |
57.00 |
56.00 |
43.00 |
40.00 |
26.00 |
30.00 |
49.00 |
43.00 |
29.00 |
38.00 |
40.80 |
41.40 |
|
Femmes |
||||||||||||
|
Arcs plats |
5.00 |
5.00 |
6.00 |
8.00 |
3.00 |
7.00 |
5.00 |
2.00 |
6.00 |
4.00 |
5.00 |
5.20 |
|
Arcs en tente |
1.00 |
0.00 |
0.00 |
3.00 |
2.00 |
6.00 |
0.00 |
4.00 |
0.00 |
2.00 |
0.60 |
3.00 |
|
Total arcs |
6.00 |
5.00 |
6.00 |
11.00 |
5.00 |
13.00 |
5.00 |
6.00 |
6.00 |
6.00 |
5.60 |
8.20 |
|
Boucles ulnaires |
50.00 |
38.00 |
53.00 |
43.00 |
72.00 |
61.00 |
57.00 |
49.00 |
63.00 |
56.00 |
59.00 |
49.40 |
|
Boucles radiales |
0.00 |
0.00 |
3.00 |
6.00 |
0.00 |
2.00 |
0.00 |
2.00 |
0.00 |
0.00 |
0.60 |
2.00 |
|
Doubles boucles |
12.00 |
21.00 |
3.00 |
6.00 |
3.00 |
2.00 |
1.00 |
0.00 |
6.00 |
33.00 |
5.00 |
12.40 |
|
Tourbillons |
32.00 |
36.00 |
35.00 |
34.00 |
20.00 |
22.00 |
37.00 |
43.00 |
25.00 |
5.00 |
29.80 |
28.00 |
|
Total tourbillons |
44.00 |
57.00 |
38.00 |
40.00 |
23.00 |
24.00 |
38.00 |
43.00 |
31.00 |
38.00 |
34.80 |
40.40 |
Tableau 2. Fréquences des figures Dermatoglyphiques par
doigt chez la population de Sabra.
Table
2.
Frequencies
figures dermatoglyphics of each fingers in the population
of Sabra.
Selon les résultats des comparaisons bimanuelles nous avons distingué une asymétrie chez les femmes pour les Boucles ulnaires et les Tourbillons entre la paire de doigts I. celle-ci pourrait être à l’origine de la différence bimanuelle constatée chez les femmes de notre population (tableau 3).
Concernant les comparaisons sexuelles, les résultats ont indiqués qu’il existe un dimorphisme sexuel pour les mains droites. Ce ci semble être lié aux différences enregistrées entre les doigts D I et D II.
|
Figures |
DI – GI |
DII – GII |
DIII–GIII |
DIV–GIV |
DV – GV |
D – G |
|
Hommes |
||||||
|
Arcs |
2.00 (NS) |
1.00 (NS) |
0.00 (NS) |
0.00 (NS) |
0.00 (NS) |
0.60 (NS) |
|
B.radiales |
2.00 (NS) |
-5.00 (NS) |
0.00 (NS) |
-1.00 (NS) |
0.00 (NS) |
-0.80 (NS) |
|
B.ulnaires |
-4.00 (NS) |
1.00 (NS) |
4.00 (NS) |
-5.00 (NS) |
9.00 (NS) |
0.80 (NS) |
|
Tourbillons |
1.00 (NS) |
3.00 (NS) |
-4.00 (NS) |
6.00 (NS) |
-9.00 (NS) |
-0.60 (NS) |
|
χ2 (signif.) |
3.317 (NS) |
1.478 (NS) |
0.424 (NS) |
0.982 (NS) |
1.901 (NS) |
0.708 (NS) |
|
ddl |
3 |
3 |
3 |
3 |
3 |
3 |
|
Femmes |
||||||
|
Arcs |
1.00 (NS) |
-5.00 (NS) |
-8.00 (NS) |
-1.00 (NS) |
0.00 (NS) |
-2.60 (NS) |
|
B.radiales |
0.00 (NS) |
-3.00 (NS) |
-2.00 (NS) |
-2.00 (NS) |
0.00 (NS) |
-1.40 (NS) |
|
B.ulnaires |
12.00 (*) |
10.00 (NS) |
11.00 (NS) |
8.00 (NS) |
7.00 (NS) |
9.60 (**) |
|
Tourbillons |
-13.00 (*) |
-2.00 (NS) |
-1.00 (NS) |
-5.00 (NS) |
-7.00 (NS) |
-5.60 (NS) |
|
χ2 (signif.) |
3.401 (NS) |
3.564 (NS) |
6.487 (NS) |
3.003 (NS) |
1.122 (NS) |
12.555 (**) |
|
ddl |
2 |
3 |
3 |
3 |
2 |
3 |
Tableau 3. Différences bimanuelles ( Droite –
Gauche ) de la fréquence des Dermatoglyphes.
D:droit, G:gauche, nombres I, II, III, IV, V:
relatifs aux doigts, χ²: test du khideux, ddl: degré de liberté.
Table
3.
Bimanuel differences (right-left) of the frequency dermatoglyphics.
D: right; G: left, numbers I, II, III, IV, V: fingers, χ²: khisquare, ddl: degree of
freedom,
Quant aux comparaisons inter- populationnelles à l’échelle de la Méditerranée, les résultats obtenus ont révélées une faible fréquence des Boucles radiales et une fréquence élevée des Tourbillons, ce qui fait rapprocher la population de Sabra en particulier à celles d’Afrique du Nord et en général à celles du Moyen Orient. Cependant, la fréquence élevée des Arcs rapproche d’avantage notre population à celles du Nord de la Méditerrané (tableau 4).
|
Sabra |
Nord de l’Afrique |
Moyen Orient |
Nord de la Méditerranée |
|
Hommes Arcs 7.5 Boucles radiales 3.0 Boucles ulnaires 48.4 Tourbillons 41.1 Femmes Arcs 9.6 Boucles radiales 1.3 Boucles ulnaires 54.2 Tourbillons 37.6 |
2.3 – 8.8 1.6 – 4.3 51.8 – 58.0 31.7 – 42.7 2.5 - 7.2 1.2 - 3.4 56.3 - 64.8 27.7 - 36.0 |
2.9 - 3.8 2.6 - 3.4 52.3 - 54.5 39.1 - 41.4 5.9 3.3 54.4 36.4 |
1.8 - 7.0 3.5 - 5.9 52.1 - 65.2 26.9 - 37.8 2.1 - 8.7 2.6 - 4.6 55.1 - 65.9 21.1 - 38.0 |
Tableau 4. Variation des fréquence des figures
dermatoglyphiques dans la population de Sabra.
Table
4.
Variation of frequencies
figures dermatoglyphics in the population of Sabra.
Résultats des groupes
sanguins
La répartition phénotypique des cinq systèmes sanguins étudiés au sein de la population de Sabra ainsi que l’estimation des fréquences géniques sont présentées dans le tableau 5. Globalement, l’hypothèse d’équilibre Hardy Weinberg est acceptée à un niveau de 1%o, pour cela nous considérons les cinq systèmes: ABO, Rhésus, MNSs, Duffy et Kell en équilibre génétique.
|
Systèmes |
Phénotypes |
Fréquences observées |
Fréquences théoriques |
Fréquences alléliques ou haplotipiques et équilibre de H.W. |
|
ABO H=0.512 |
A1 A2 B A1B A2B O |
48 21 31 8 4 91 |
47.312 19.708 31.072 6.714 2.969 90.855 |
ABO * A1 = 0.156 ABO * A2 = 0.069 ABO * B = 0.106 ABO * O = 0.669 χ2
HW (1) = 0.699 (NS) |
|
Total = 203 |
||||
|
Rhésus H=0.374 |
CCD-EE CCD-Ee CCD-ee CcD-EE CcD-Ee CcD-ee ccD-EE ccD-Ee ccD-ee CCddEE CCddEe CCddee CcddEE CcddEe Ccddee ccddEE ccddEe ccddee |
0 1 17 0 12 36 3 13 12 0 0 0 0 0 0 0 0 6 |
0 1.022 16.894 0 12.267 36.540 2.560 12.862 11.925 0 0 0 0 0 0 0 0 5.953 |
Rh * D = 0.756 Rh * d = 0.244 Rh * C = 0.420 Rh * c = 0.580 Rh * E = 0.160 Rh * e = 0.840 Rh * CDE = 0.000 Rh * CDe = 0.415 Rh * cDE = 0.155 Rh * cDe = 0.186 Rh * CdE = 0.000 Rh * Cde = 0.000 Rh * cdE = 0.000 Rh * cde = 0.244 χ2 HW (1) = 0.092 (NS) |
|
Total = 100 |
||||
|
MNSs H=0.410 |
MMSS MMSs MMss MNSS MNSs MNss NNSS NNSs NNss |
1 3 2 1 1 11 0 0 3 |
0.480 3.560 2.373 0.830 0.830 9.460 0 0 4.080 |
MN * M = 0.569 MN * N = 0.431 Ss * S = 0.209 Ss * s = 0.791 MNSs * MS = 0.144 MNSs * Ms = 0.421 MNSs * NS = 0.036 MNSs * Ns = 0.399 χ2 HW (1) = 1.316 (NS) |
|
Total = 22 |
||||
|
Duffy H=0.641 |
Fy (a+, b+) Fy (a+, b-) Fy (a-, b+) Fy (a-, b-) |
6 6 9 1 |
6.539 5.702 8.683 1.074 |
Fy * A = 0.334 Fy * B = 0.445 Fy * O = 0.221 χ2 HW (1) = 0.076 (NS) |
|
Total = 22 |
||||
|
Kell H=0.095 |
K+ K- |
9 91 |
9.75 90.25 |
Kell * K = 0.050 Kell * k = 0.950 χ2 HW (1) = 0.058 (NS) |
|
Total = 100 |
||||
Tableau 5. Fréquences alléliques et haplotypiques des
systèmes sanguins (ABO, Rhésus, MNSs, Duffy et Kell) dans la population de
Sabra.
Table
5.
allelic and haplotypic frequencies of blood system (ABO, Rhésus, MNSs, Duffy
and Kell) in population of Sabra.
Système ABO
Nous retrouvons une prédominance de l’allèle O (66.9%) par rapport aux allèles A (22.5%) et B (10.6%). Par ailleurs, le taux de l’allèle A2 reste faible (6.9%) si l’on compare avec celui de l’allèle A1 (15.6%).
En ce qui concerne, la comparaison de ces fréquences alléliques de la population de Sabra par rapport à celles enregistrées en Algérie tableau 6, elle évoque une grande similitude avec de nombreuses populations. Cette similitude est détectable avec l’ensemble des allèles.
Quant à la comparaison des fréquences géniques ABO de la population de Sabra part rapport à celles de l’Algérie, elle montre une grande homogénéité avec un pourcentage de différences non significatives (%DNS) estimé à 92.30%. Cependant, les différences significatives ont été retrouvées avec la wilaya de Mostaganem (MOS) et les Oasis (OAS), ce ci pourrait être du aux fréquences élevées de l’allèle ABO*B à Mostaganem ainsi de même de l’allèle ABO*O dans les Oasis.
|
Populations des wilayas |
N |
ABO*A |
ABO*B |
ABO*O |
χ2 |
Références |
|
Sabra |
203 |
0.225 |
0.106 |
0.669 |
|
Présente étude |
|
Alger |
347 |
0.209 |
0.120 |
0.671 |
0.696 (NS) |
Aireche et Benabadji, 1994 |
|
Annaba |
126 |
0.247 |
0.126 |
0.627 |
1.366 (NS) |
|
|
Batna |
160 |
0.230 |
0.085 |
0.685 |
0.956 (NS) |
|
|
Bejaia |
176 |
0.208 |
0.135 |
0.657 |
1.754 (NS) |
|
|
Biskra |
98 |
0.218 |
0.125 |
0.657 |
0.589 (NS) |
|
|
Blida |
192 |
0.273 |
0.122 |
0.605 |
3.743 (NS) |
|
|
Boumerdes |
178 |
0.213 |
0.135 |
0.652 |
1.522 (NS) |
|
|
Chlef |
204 |
0.259 |
0.112 |
0.629 |
1.659 (NS) |
|
|
Constantine |
222 |
0.217 |
0.115 |
0.668 |
0.215 (NS) |
|
|
Guelma |
264 |
0.184 |
0.112 |
0.704 |
2.333 (NS) |
|
|
Jijel |
170 |
0.183 |
0.123 |
0.694 |
2.238 (NS) |
|
|
Mascara |
95 |
0.227 |
0.130 |
0.643 |
0.902 (NS) |
|
|
Media |
111 |
0.185 |
0.105 |
0.710 |
1.436 (NS) |
|
|
Mostaganem |
127 |
0.171 |
0.161 |
0.668 |
6.097 (*) |
|
|
M’Sila |
85 |
0.180 |
0.099 |
0.721 |
1.530 (NS) |
|
|
Oum el Bouaki |
183 |
0.200 |
0.132 |
0.668 |
1.595 (NS) |
|
|
Oran |
89 |
0.219 |
0.157 |
0.624 |
3.108 (NS) |
|
|
Saida |
63 |
0.242 |
0.117 |
0.641 |
0.341 (NS) |
|
|
Setif |
339 |
0.189 |
0.138 |
0.673 |
3.726 (NS) |
|
|
Sidi bel Abbes |
113 |
0.208 |
0.088 |
0.704 |
0.856 (NS) |
|
|
Skikda |
149 |
0.219 |
0.145 |
0.636 |
2.375 (NS) |
|
|
Tebessa |
126 |
0.185 |
0.124 |
0.691 |
1.551 (NS) |
|
|
Tiaret |
119 |
0.166 |
0.151 |
0.683 |
4.884 (NS) |
|
|
Tizi-Ouzou |
475 |
0.243 |
0.120 |
0.637 |
1.403 (NS) |
|
|
Tlemcen |
139 |
0.159 |
0.102 |
0.739 |
4.747 (NS) |
|
|
Oasis |
94 |
0.132 |
0.131 |
0.737 |
7.185 (*) |
|
|
Total |
4444 |
0.209 |
0.123 |
0.668 |
|
Tableau 6. Comparaisons de la distribution des
fréquences alléliques du système ABO de la population de Sabra avec celles des
populations d’Algérie.
Table
6.
Comparison of the distribution of allele frequencies of the ABO system of
population Sabra with those of populations of Algeria.
Système Rhésus
Il est représenté par 94% d’individus de phénotypes Rh+ et 6% de phénotype Rh-. L’haplotype CDe apparaît comme le plus fréquent avec une fréquence de 41.5%. Cet haplotype avec trois autres qui sont cde, cDe et cDE représentent les haplotypes les plus communs dans la population de Sabra. Parmi les haplotypes considérés comme exceptionnelles et rares CDE, CdE, Cde et cdE, aucun n’a été mis en évidence comme cela est souvent le cas dans la majorité des populations.
Quant aux comparaisons de la distribution des fréquences haplotypiques avec celles d’Algérie mentionnées dans le tableau 7, l’analyse met en évidence une certaine hétérogénéité de la répartition haplotypique du système Rhésus de la population de Sabra par rapport aux populations Algériennes,
L’haplotype cDE avec une fréquence de près de 15.5% dépasse largement celle de la moyenne Algérienne (9.5%). Ce ci nous laisse supposer qu’il est à l’origine de la variation du système Rhésus chez la population de Sabra.
D’autres parts, les résultats de comparaison démontrent aussi à l’échelle de l’Algérie un pourcentage de différences non significatives estimé à 53.85%. Les différences significatives ont été trouvées avec les wilayas de BAT, BEJ, BLI, BOU, GUE, MOS, SAI, SAB, SKI, TOU, TLE et les OAS.
|
Populations |
N |
CDE |
CDe |
cDE |
cDe |
CdE |
Cde |
cdE |
cde |
χ2 |
Références |
|
Sabra |
100 |
0.000 |
0.415 |
0.155 |
0.186 |
0.000 |
0.000 |
0.000 |
0.244 |
|
Présente étude |
|
Alger |
315 |
0.000 |
0.442 |
0.098 |
0.198 |
0.000 |
0.016 |
0.000 |
0.252 |
7.927 (NS) |
Aireche et Benabadji, 1988. |
|
Annaba |
135 |
0.000 |
0.456 |
0.082 |
0.206 |
0.000 |
0.000 |
0.000 |
0.256 |
6.281 (NS) |
|
|
Batna |
155 |
0.000 |
0.458 |
0.119 |
0.263 |
0.000 |
0.019 |
0.000 |
0.141 |
15.916 (*) |
|
|
Bejaia |
164 |
0.000 |
0.461 |
0.073 |
0.184 |
0.000 |
0.021 |
0.000 |
0.261 |
12.978 (*) |
|
|
Biskra |
91 |
0.000 |
0.356 |
0.120 |
0.202 |
0.000 |
0.018 |
0.000 |
0.304 |
6.229 (NS) |
|
|
Blida |
172 |
0.000 |
0.420 |
0.076 |
0.283 |
0.000 |
0.026 |
0.014 |
0.181 |
23.185(***) |
|
|
Boumerdes |
186 |
0.000 |
0.413 |
0.065 |
0.201 |
0.000 |
0.017 |
0.000 |
0.304 |
16.071 (**) |
|
|
Chlef |
199 |
0.000 |
0.364 |
0.113 |
0.255 |
0.000 |
0.021 |
0.000 |
0.247 |
9.487 (NS) |
|
|
Constantine |
220 |
0.000 |
0.448 |
0.096 |
0.189 |
0.000 |
0.000 |
0.000 |
0.277 |
5.157 (NS) |
|
|
Guelma |
262 |
0.000 |
0.393 |
0.116 |
0.255 |
0.000 |
0.035 |
0.000 |
0.201 |
13.160 (*) |
|
|
Jijel |
168 |
0.000 |
0.420 |
0.098 |
0.251 |
0.000 |
0.000 |
0.000 |
0.231 |
5.826 (NS) |
|
|
Mascara |
96 |
0.000 |
0.339 |
0.104 |
0.236 |
0.000 |
0.000 |
0.000 |
0.321 |
6.704 (NS) |
|
|
Media |
104 |
0.000 |
0.360 |
0.115 |
0.218 |
0.000 |
0.025 |
0.000 |
0.282 |
7.849 (NS) |
|
|
Mostaganem |
127 |
0.000 |
0.394 |
0.079 |
0.272 |
0.000 |
0.016 |
0.000 |
0.239 |
12.678 (*) |
|
|
M’Sila |
78 |
0.000 |
0.532 |
0.109 |
0.172 |
0.000 |
0.000 |
0.000 |
0.187 |
5.419 (NS) |
|
|
Oum el Bouaki |
184 |
0.000 |
0.473 |
0.098 |
0.212 |
0.000 |
0.000 |
0.000 |
0.217 |
5.448 (NS) |
|
|
Oran |
88 |
0.000 |
0.352 |
0.114 |
0.240 |
0.000 |
0.018 |
0.000 |
0.276 |
7.228 (NS) |
|
|
Saida |
62 |
0.000 |
0.358 |
0.105 |
0.239 |
0.000 |
0.030 |
0.000 |
0.268 |
9.911 (*) |
|
|
Setif |
333 |
0.000 |
0.460 |
0.102 |
0.207 |
0.000 |
0.007 |
0.000 |
0.224 |
6.623 (NS) |
|
|
Sidi bel Abbes |
112 |
0.012 |
0.366 |
0.058 |
0.231 |
0.000 |
0.025 |
0.036 |
0.272 |
26.824(***) |
|
|
Skikda |
148 |
0.000 |
0.453 |
0.087 |
0.186 |
0.000 |
0.027 |
0.000 |
0.247 |
10.480 (*) |
|
|
Tebessa |
125 |
0.010 |
0.442 |
0.086 |
0.209 |
0.000 |
0.000 |
0.000 |
0.253 |
6.371 (NS) |
|
|
Tiaret |
114 |
0.000 |
0.393 |
0.092 |
0.240 |
0.000 |
0.016 |
0.000 |
0.259 |
8.733 (NS) |
|
|
Tizi-Ouzou |
455 |
0.002 |
0.434 |
0.083 |
0.183 |
0.000 |
0.018 |
0.004 |
0.276 |
14.244 (*) |
|
|
Tlemcen |
137 |
0.008 |
0.414 |
0.065 |
0.211 |
0.000 |
0.023 |
0.000 |
0.279 |
15.631 (**) |
|
|
Oasis |
91 |
0.000 |
0.361 |
0.139 |
0.338 |
0.000 |
0.000 |
0.000 |
0.162 |
13.205 (**) |
|
|
Total |
4321 |
0.001 |
0.421 |
0.095 |
0.218 |
0.000 |
0.015 |
0.002 |
0.248 |
|
Tableau 7. Comparaisons de la distribution des
fréquences haplotipiques du système Rhésus de la population de Sabra avec
celles des populations des wilayas d’Algérie.
Table
7.
Comparisons of the frequency distribution of Rhesus haplotipiques population
Sabra with those populations wilaya of Algeria.
Système MNSs
Les résultats concernant la comparaison de la répartition des fréquences haplotypiques de ce système MNSs chez la population de Sabra avec les populations d’Algérie sont indiqués dans le tableau 8. Nous constatons que l’allèle M (56.9%) dépasse proportionnellement l’allèle N (43.1%), alors que l’allèle S (20.9%) est largement en dessous de l’allèle s (79.1%).
Dans l’ensemble, l’analyse ne démontre pas de grande variabilité à l’échelle nationale
En fait, l’haplotype le plus répandu dans la distribution de notre population est MNSs*Ms (42.1%). Cette fréquence dépasse fréquemment celle de la moyenne Algérienne (30.8%)
L’haplotype MNSs*Ns marque une fréquence de 39.9%, qui est comprise dans l’intervalle de variations des populations d’Algérie (31.8%-50.8%).
Par ailleurs, l’haplotype MNSs*MS est observé avec une fréquence de 14.4%. Celle-ci se situe au milieu de l’intervalle de variations des Algériennes (15.6%-23.6%).
Néanmoins, l’haplotype MNSs*NS apparaît avec une fréquence relativement faible de 3.6% par rapport à celles des populations analysées.
Globalement, pour ce système, les résultats révèlent une homogénéité de notre population comparée aux populations Algériennes, une seule différence a été marquée avec la population de la wilaya de JIJ, le %DNS est évalué à 96.15%.
|
Populations des wilayas |
N |
MS |
Ms |
NS |
Ns |
χ2 |
Références |
|
Sabra |
22 |
0.144 |
0.421 |
0.036 |
0.399 |
|
Présente étude. |
|
Alger |
338 |
0.201 |
0.280 |
0.099 |
0.420 |
5.447 (NS) |
Aireche et Benabadji, 1990. |
|
Annaba |
135 |
0.192 |
0.272 |
0.082 |
0.454 |
4.838 (NS) |
|
|
Batna |
158 |
0.196 |
0.377 |
0.089 |
0.338 |
2.455 (NS) |
|
|
Bejaia |
167 |
0.178 |
0.266 |
0.124 |
0.432 |
6.174 (NS) |
|
|
Biskra |
96 |
0.156 |
0.355 |
0.058 |
0.431 |
0.814 (NS) |
|
|
Blida |
188 |
0.221 |
0.288 |
0.058 |
0.433 |
4.219 (NS) |
|
|
Boumerdes |
186 |
0.165 |
0.303 |
0.117 |
0.415 |
4.109 (NS) |
|
|
Chlef |
209 |
0.188 |
0.305 |
0.068 |
0.439 |
3.035 (NS) |
|
|
Constantine |
222 |
0.183 |
0.344 |
0.066 |
0.407 |
1.589 (NS) |
|
|
Guelma |
260 |
0.211 |
0.313 |
0.048 |
0.428 |
2.919 (NS) |
|
|
Jijel |
165 |
0.233 |
0.233 |
0.054 |
0.48 |
8.064 (*) |
|
|
Mascara |
96 |
0.207 |
0.319 |
0.053 |
0.421 |
2.371 (NS) |
|
|
Media |
109 |
0.236 |
0.352 |
0.094 |
0.318 |
4.099 (NS) |
|
|
Mostaganem |
127 |
0.179 |
0.341 |
0.081 |
0.399 |
1.891 (NS) |
|
|
M’Sila |
84 |
0.186 |
0.343 |
0.058 |
0.413 |
1.305 (NS) |
|
|
Oum el Bouaki |
161 |
0.158 |
0.290 |
0.049 |
0.503 |
3.417 (NS) |
|
|
Oran |
86 |
0.218 |
0.335 |
0.090 |
0.357 |
3.031 (NS) |
|
|
Saida |
62 |
0.217 |
0.324 |
0.082 |
0.377 |
2.837 (NS) |
|
|
Setif |
343 |
0.182 |
0.359 |
0.074 |
0.385 |
1.633 (NS) |
|
|
Sidi bel Abbes |
114 |
0.217 |
0.345 |
0.077 |
0.361 |
2.679 (NS) |
|
|
Skikda |
145 |
0.229 |
0.303 |
0.036 |
0.432 |
3.611 (NS) |
|
|
Tebessa |
123 |
0.162 |
0.277 |
0.053 |
0.508 |
3.948 (NS) |
|
|
Tiaret |
117 |
0.232 |
0.298 |
0.093 |
0.377 |
4.683 (NS) |
|
|
Tizi-Ouzou |
467 |
0.173 |
0.293 |
0.103 |
0.431 |
4.452 (NS) |
|
|
Tlemcen |
136 |
0.232 |
0.287 |
0.073 |
0.408 |
4.599 (NS) |
|
|
Oasis |
94 |
0.177 |
0.394 |
0.078 |
0.351 |
1.360 (NS) |
|
|
Total |
4388 |
0.194 |
0.308 |
0.078 |
0.420 |
|
Tableau 8. Comparaisons de la distribution des fréquences
haplotipiques du système MNSs de la population de Sabra avec celles des
populations des wilayas d’Algérie.
Table
8.
Comparison of frequency distribution haplotipiques MNSs system of population
with those populations Sabra wilaya of Algeria.
Système Duffy
L’allèle apparaissant le plus fréquent à Sabra est Fy*b avec une fréquence de (44.5%) suivi de l’allèle Fy*a (33.4%) et en dernier lieu l’allèle Fy*o (22.1%).
Pour la comparaison des fréquences alléliques concernant ce système, les résultats mentionnés dans le tableau 9, montrent que notre population présente des fréquences géniques des trois allèles comparables à celles trouvées dans les populations Algériennes à l’exception d’ANA, BEJ et les OAS ou nous avons remarqué une différence, le %DNS est estimé à 88.46%.
|
Populations des wilayas |
N |
Fy * a |
Fy * b |
Fy * o |
χ2 |
Références |
|
Sabra |
22 |
0.334 |
0.445 |
0.221 |
|
Présente étude. |
|
Alger |
295 |
0.269 |
0.445 |
0.286 |
1.276 (NS) |
Aireche et Benabadji, 1988. |
|
Annaba |
137 |
0.199 |
0.337 |
0.464 |
9.814 (**) |
|
|
Batna |
160 |
0.292 |
0.514 |
0.194 |
0.806 (NS) |
|
|
Bejaia |
175 |
0.276 |
0.611 |
0.113 |
6.111 (*) |
|
|
Biskra |
95 |
0.224 |
0.390 |
0.386 |
4.750 (NS) |
|
|
Blida |
189 |
0.271 |
0.479 |
0.250 |
0.858 (NS) |
|
|
Boumerdes |
179 |
0.244 |
0.518 |
0.238 |
1.794 (NS) |
|
|
Chlef |
209 |
0.244 |
0.476 |
0.280 |
1.916 (NS) |
|
|
Constantine |
223 |
0.244 |
0.454 |
0.302 |
2.244 (NS) |
|
|
Guelma |
265 |
0.231 |
0.512 |
0.257 |
2.482 (NS) |
|
|
Jijel |
169 |
0.279 |
0.543 |
0.178 |
1.652 (NS) |
|
|
Mascara |
96 |
0.256 |
0.478 |
0.266 |
1.219 (NS) |
|
|
Media |
111 |
0.287 |
0.397 |
0.316 |
1.595 (NS) |
|
|
Mostaganem |
126 |
0.245 |
0.495 |
0.260 |
1.603 (NS) |
|
|
M’Sila |
84 |
0.291 |
0.451 |
0.258 |
0.395 (NS) |
|
|
Oum el Bouaki |
183 |
0.277 |
0.500 |
0.223 |
0.751 (NS) |
|
|
Oran |
87 |
0.298 |
0.412 |
0.290 |
0.801 (NS) |
|
|
Saida |
63 |
0.337 |
0.471 |
0.192 |
0.240 (NS) |
|
|
Setif |
344 |
0.336 |
0.516 |
0.148 |
1.996 (NS) |
|
|
Sidi bel Abbes |
115 |
0.227 |
0.485 |
0.288 |
2.546 (NS) |
|
|
Skikda |
150 |
0.229 |
0.507 |
0.264 |
2.334 (NS) |
|
|
Tebessa |
127 |
0.330 |
0.515 |
0.155 |
1.529 (NS) |
|
|
Tiaret |
117 |
0.259 |
0.423 |
0.318 |
1.928 (NS) |
|
|
Tizi-Ouzou |
467 |
0.340 |
0.513 |
0.147 |
2.096 (NS) |
|
|
Tlemcen |
137 |
0.321 |
0.437 |
0.242 |
0.099 (NS) |
|
|
Oasis |
94 |
0.239 |
0.305 |
0.456 |
8.448 (*) |
|
|
Total |
4397 |
0.274 |
0.475 |
0.251 |
|
Tableau 9. Comparaisons de la distribution des
fréquences alléliques (Fy * a, Fy * b et Fy * o) du système Duffy de la
population de Sabra avec celles des populations des wilayas d’Algérie.
Table
9.
Comparison of the distribution of allele frequencies (Fy * a, b and Fy * Fy *
o) of the Duffy system of population with those populations Sabra wilaya of
Algeria.
Système Kell
Nous notons une faible fréquence de l’allèle Kell*K (5%) par rapport à l’allèle Kell*k (95%).
D’après les résultats de la comparaison de la répartition des fréquences alléliques du système Kell présentés dans le tableau 10, notre population apparaît avec des valeurs très similaires à celles observées dans toutes les populations des wilayas d’Algérie. Le %DNS est estimé à 100%.
|
Populations des wilayas |
N |
Kell * K |
Kell * k |
χ2 |
Références |
|
Sabra |
100 |
0.050 |
0.950 |
|
Présente étude. |
|
Alger |
302 |
0.041 |
0.959 |
0.267 (NS) |
Aireche et Benabadji, 1995. |
|
Annaba |
93 |
0.054 |
0.946 |
0.028 (NS) |
|
|
Batna |
128 |
0.027 |
0.973 |
1.606 (NS) |
|
|
Bejaia |
143 |
0.063 |
0.937 |
0.363 (NS) |
|
|
Biskra |
76 |
0.059 |
0.941 |
0.143 (NS) |
|
|
Blida |
145 |
0.028 |
0.972 |
1.681 (NS) |
|
|
Boumerdes |
170 |
0.047 |
0.953 |
0.024 (NS) |
|
|
Chlef |
170 |
0.059 |
0.941 |
0.187 (NS) |
|
|
Constantine |
176 |
0.043 |
0.957 |
0.161 (NS) |
|
|
Guelma |
199 |
0.068 |
0.932 |
0.730 (NS) |
|
|
Jijel |
146 |
0.034 |
0.966 |
0.755 (NS) |
|
|
Mascara |
71 |
0.049 |
0.951 |
0.001 (NS) |
|
|
Media |
79 |
0.063 |
0.937 |
0.296 (NS) |
|
|
Mostaganem |
94 |
0.048 |
0.952 |
0.009 (NS) |
|
|
M’Sila |
62 |
0.065 |
0.935 |
0.307 (NS) |
|
|
Oum el Bouaki |
142 |
0.039 |
0.961 |
0.359 (NS) |
|
|
Oran |
63 |
0.056 |
0.944 |
0.048 (NS) |
|
|
Saida |
49 |
0.102 |
0.898 |
2.845 (NS) |
|
|
Setif |
290 |
0.055 |
0.945 |
0.078 (NS) |
|
|
Sidi bel Abbes |
85 |
0.053 |
0.947 |
0.016 (NS) |
|
|
Skikda |
107 |
0.042 |
0.958 |
0.149 (NS) |
|
|
Tebessa |
102 |
0.049 |
0.951 |
0.002 (NS) |
|
|
Tiaret |
93 |
0.027 |
0.973 |
1.379 (NS) |
|
|
Tizi-Ouzou |
421 |
0.057 |
0.943 |
0.151 (NS) |
|
|
Tlemcen |
110 |
0.032 |
0.968 |
0.892 (NS) |
|
|
Oasis |
74 |
0.034 |
0.966 |
0.542 (NS) |
|
|
Total |
3590 |
0.049 |
0.951 |
|
Tableau 10. Comparaisons de la distribution des
fréquences alléliques du système Kell de la population de Sabra avec celles des
populations des wilayas d’Algérie.
Table
10.
Comparison of the distribution of allele frequencies of Kell system of
population with those populations Sabra wilaya of Algeria.
Affinités inter-
populationnelles
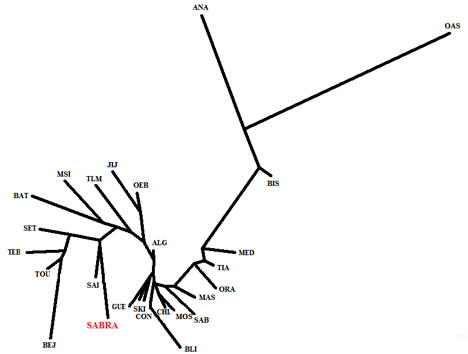
Au total, deux types d’analyses: en composantes principales et en fonction des distances génétiques outre leurs représentation sous forme d’arbres phylogénétiques ont été consacrées pour la mise en évidence des affinités inter- populationnelles.
En Algérie, pour l’analyse en composantes principales, les résultats sont représentés en figure 2. Les deux premiers axes présentent respectivement 18.70% et 14.90%, soit un total de 33.60% de la variabilité globale. Nous constatons que la répartition des populations des wilayas d’Algérie en fonction des systèmes sanguins est plus ou moins corrélée à leurs dispositions géographiques. Selon les deux axes la population de Sabra occupe une position centrale au sein de la figure dans les deux cotés des abscisses et des ordonnées positifs. De plus, ce schéma confirme aussi le regroupement de notre population avec toutes les populations des différentes wilayas d’Algérie mis à part celles de JIJ, OEB, TEB, TOU, SAI et les OAS où elles apparaissent très différenciées. Ce ci semble être lié aux fréquences géniques caractéristiques de chacune de ces wilayas.
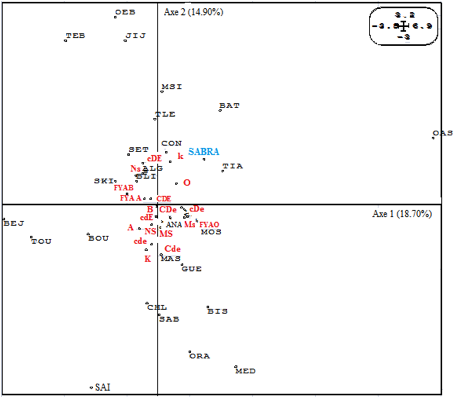
Figure 2. Analyse en composantes principales de la
distribution des systèmes sanguins à l’échelle de l'Algérie.
Figure 2. Principal component analysis of the distribution of blood
systems across Algeria.
Cependant, les distances génétiques calculées d’après la formule de Renolds, attribuent aux populations D’Algérie une distance génétique Moyenne de (109×10-4) (tableau 11, 12).