
Sanna, E., Iovine, M. C., Calò, C. M., 2006, La deriva genetica ed il flusso genico interno hanno condizionato l’attuale struttura biologica della popolazione sarda? Antropo, 12, 43-52. www.didac.ehu.es/antropo
La deriva genetica ed il flusso genico interno hanno condizionato l’attuale struttura biologica della popolazione sarda?
Did genetic drift and gene flow influence the present biological structure of Sardinian population?
Emanuele Sanna, Maria Chiara Iovine, Carla Maria Calò
Dipartimento di Biologia Sperimentale, Sezione di Scienze Antropologiche, Università degli Studi di Cagliari, Italia
Per la corrispondenza: Emanuele Sanna, Dipartimento di Biologia Sperimentale, Sezione di Scienze Antropologiche, Università degli Studi di Cagliari, Cittadella Universitaria, SS 554 (km 4,5), 09042 Monserrato, Italia. E-mail: sanane@unica.it
Parole Chiave: Sardegna, mtDNA, cromosoma Y, isolamento, collo di bottiglia,
Key words: Sardinia, mtDNA, Y chromosome, isolation, bottleneck
Riassunto
La specificità biologica della popolazione sarda nel contesto delle popolazioni europee e circum-mediterranee nonché la sua eterogeneità interna troverebbe supporto in conseguenza dell'azione di diversi fattori microevolutivi che hanno agito nel tempo (effetto del fondatore, effetto collo di bottiglia, isolamento e flussi genici interni). Inoltre anche la peculiarità biologica della popolazione sarda stanziata nella cosiddetta “Area arcaica” dell’Isola potrebbe essere una conseguenza dell’azione della deriva genetica e dell’isolamento. Sostiene questa ipotesi la più elevata incidenza nell’Isola, sinora riscontrata, della mutazione M26 del cromosoma Y (33-41%) rispetto alle altre popolazioni dell’Europa continentale e dell’area mediterranea, probabile conseguenza della diffusione di gruppi umani portatori di tale mutazione, distaccatisi dalla popolazione ancestrale continentale dopo Last Glacial Maximum. L’eterogeneità tra le sottopopolazioni della Sardegna trova supporto nei numerosi decrementi di popolazione, soprattutto a carico di quelle delle aree interne dell’Isola, deducibili dai dati di demografia storica relativi ai Censimenti fiscali effettuati nel Trecento durante la dominazione pisana prima ed a quelli spagnoli successivamente nel Seicento. In sintesi i dati di demografia storica consentono di spiegare in modo congruente l’attuale struttura biologica della popolazione sarda.
Abstract
The biological peculiarity of Sardinia among the European and Mediterranean populations find a support in the consequences of the action of some micro-evolutionary factors that acted in time (founder effect, bottleneck, isolation, and internal gene flow). Moreover, the biological peculiarity of the Sardinian population located in the so-called “Archaic area” of the island could be a consequence of genetic drift and isolation actions. This hypothesis is supported by the highest incidence in the island of the mutation M26 of Y chromosome (33-41%) with respect to the other populations from Europe and Mediterranean basin. This high frequency could be due to the diffusion of men carrying that mutation, split by the continental ancestor group after the Last Maximum Glacial. The heterogeneity among the Sardinian sub-populations find a support in several decreases of the population, particularly charged to the ones of internal area of the island, deduced by historical demographic data of Census carried out in the 300 during the Pisan domination, and later by the Spaniards during the 600. In conclusion the historical demographic data allow to explain adequately the present biological structure of Sardinian population.
Introduzione
Gli studi sulle relazioni biologiche tra i Sardi e le altre popolazioni dimostrano che la popolazione della Sardegna presenta un quadro biologico peculiare che la differenzia nettamente dalle popolazioni del continente europeo e circum-mediterranee (Vona, 1995; Memmì et al., 1998; Lampis et al., 2000; Ghiani e Vona, 2002; Vona et al., 2002; Francalacci et al., 2003; Quintana-Murci et al., 2003).
Cavalli-Sforza et al. (1994) sostengono che la specificità biologica dei Sardi sia una conseguenza della deriva genetica e che l’apporto di geni da parte dei Neolitici sia stato limitato e forse con apporto di nuova deriva (Cavalli-Sforza et al., 1994).
Altri Autori (Chiarelli, 2003; Francalacci et al., 2003; Rootsi et al., 2004) ipotizzano che la peculiare elevata incidenza della mutazione M26 del cromosoma Y nei Sardi sarebbero una conseguenza dell’effetto del fondatore nell’espansione dalla popolazione ancestrale dopo L’ultimo Massimo Glaciale (Last Glacial Maximum).
Secondo Memmì et al. (1998) la
popolazione sarda e quella corsa presenterebbero, nell’ambito delle popolazioni
mediterranee, una forte similitudine nella struttura biologica dovuta
all’origine da una popolazione ancestrale comune; mentre le differenze
genetiche riscontrabili tra le due popolazioni risalirebbero non al Neolitico
ma all’azione di successivi e relativamente recenti effetti collo di bottiglia
(bottleneck) sulle due popolazioni. Una ipotesi simile è sostenuta da Latini et
al.
(2003) in base alle frequenze degli aplotipi del cluster ![]() -globinico.
-globinico.
Infine da notare che Falchi et al. (2004) e Piras et al. (2005) oltre ad aver ipotizzato la comune origine dei Sardi e dei Corsi, in base ai propri risultati ed a quanto riportato da vari Autori (Varesi et al., 2000; Vona et al., 2002; Vona et al., 2003), sostengono inoltre che le differenze riscontrabili tra le due popolazioni siano una conseguenza dell’azione di svariati fenomeni che producono la deriva genetica: la bassa densità demografica, le epidemie che possono aver causato un effetto bottleneck ed i lunghi periodi di isolamento che hanno caratterizzato diversi centri sia corsi sia sardi.
In questo lavoro s’intende: 1) discutere la possibilità che l’effetto fondatore e l’effetto bottleneck possano aver influito sulla struttura biologica dei Sardi; 2) valutare quali possano essere stati ed in quali zone storico-geografiche dell’Isola possano essere avvenuti degli episodi di crollo demografico e dunque possibili effetti bottleneck.
Materiali e metodi
Le stime della popolazione del periodo romano adottate sono quelle proposte per l'ultimo secolo della Repubblica da Pais (1923): 300.000 individui, mentre Meloni (1975) per il periodo individuabile nel I secolo avanti Cristo ne propone 150.000.
Le stime della popolazione del 1320 e del 1350 sono quelle riportate da Day (1987) in base ad estrapolazioni derivate dai Censimenti fiscali pisani o pisano-catalani (Day, 1987).
Per il confronto demografico tra i Censimenti fiscali spagnoli del 1583/1603, del 1627 e del 1655 i decrementi sono stati calcolati in base al numero dei “fuochi”. I primi Censimenti fiscali spagnoli erano finalizzati all’individuazione dei “fuochi”, cioè dei nuclei familiari tassabili per il contributo a quei “donativi” periodici, che ogni stato della Corona spagnola “offriva” al sovrano (Serri, 1997a). Il numero dei “fuochi” dei Censimenti del 1583, del 1627 e del 1655 sono quelli indicati da Serri (1997a; 1997b) per singolo villaggio dell’Isola.
I “fuochi” e la popolazione dei Censimenti fiscali spagnoli del 1678 e del 1688 sono quelli riportati da Corridore (1902) per singolo villaggio della Sardegna.
La perimetrazione delle zone storico-geografiche della Sardegna adottata con i relativi villaggi che le compongono è quella proposta da Ghiani Moi (1964).
Le zone storico-geografiche
Tra il IX ed il XII secolo la Sardegna era divisa in 4 Giudicati autonomi: di Arborea, di Cagliari, di Gallura e di Torres o di Logudoro. Il Giudice (Judike) era la suprema autorità militare e civile del rispettivo Giudicato (rennu o logu). Il Giudice governava e legiferava con l’assistenza di un’assemblea (corona de logu) costituita dai rappresentanti dei centri abitati (villas) che formavano il Giudicato. Ogni Giudicato era suddiviso in distretti territoriali minori (curatorias), ciascuno dei quali comprendeva diversi villaggi (villas), a loro volta rette da un “majore de villa” nominato annualmente dal Giudice o dal “curatore”, ufficiale regio che sovraintendeva alle attività amministrative ed all’esercizio della giustizia nella “curatoria” in nome del Giudice (Besta, 1908-1909). Questo tipo di sistema governativo rimase inalterato sino alla caduta del rispettivo Giudicato: nel 1256 terminò il Giudicato di Cagliari, nel 1259 quello di Torres, nel 1296 quello di Gallura e nel 1410 ebbe fine il Giudicato di Arborea (Caocci, 1985).
Nel periodo giudicale, unico periodo storico in cui i Sardi godettero dell’indipendenza amministrativa e politica, l’Isola venne suddivisa in una sessantina di distretti territoriali minori (curatorias) (Besta, 1908-1909; Mori, 1975; Casula, 1980; Day, 1982, 1987) che rispettavano sia l’ambiente fisico sia quello umano (Terrosu Asole, 1982).
Durante le dominazioni successive: catalano-aragonese (1323-1478), spagnola (1479-1714), austriaca (1714-1720), piemontese (1720-1861) (Caocci, 1985), l’Isola fu smembrata in numerosi feudi che non tenevano conto della costituzione geografica del territorio e delle esigenze della popolazione. Pertanto, con la scomparsa della forma politica ed organizzativa del territorio in vigore nel periodo giudicale, si perse non solo la concezione di molte regioni naturali, soprattutto di quelle geograficamente meno caratterizzate, ma caddero in disuso anche i coronimi che le identificavano. Fra i nomi regionali dei distretti territoriali minori del periodo giudicale, certi sono sopravvissuti inglobando aree appartenenti a curatorie limitrofe, alcuni hanno mutato dizione, altri infine hanno continuato ad indicare le medesime aree (Terrosu Asole, 1980, 1982). Nell’uso comune sono sopravvissuti una trentina di nominativi che identificano altrettante zone storico-geografiche dell’Isola, cioè quelle meglio caratterizzate dai fattori geografici (Ghiani Moi, 1964; Mori, 1975; Day, 1987).
Risultati e discussione
In Tabella 1 sono riportate le variazioni percentuali dei “fuochi” nelle zone storico-geografiche della Sardegna tra i Censimenti del 1583/1603, 1627, 1655 e della popolazione tra il Censimento del 1678 e quello del 1688.
Effetto del fondatore (Founder effect)
L’azione dell’effetto del fondatore sulla struttura genetica di una comunità sarda è stata studiata da Angius et al. (2001) che hanno evidenziato, in base ad aplotipi del cromosoma Y ed aplogruppi del DNA mitocondriale, la peculiarità biologica della comunità di Talana (Ogliastra). In questo studio che si basa sull’analisi di dati demografici, biodemografici e genetici è stato accertato che circa l’80% dell’attuale popolazione del Comune deriverebbe da otto ancestrali linee paterne ed undici materne (Angius et al., 2001, p. 198). I risultati di questo lavoro rimarcano dunque come l’effetto del fondatore ed il successivo isolamento possano influire sulla struttura genetica anche di singole comunità dell’Isola.
Falchi et al. (2005) avendo
riscontrato un’elevata incidenza della mutazione ![]() 039 nel centro della
Corsica (88,40%) ed in Sardegna (95,40%), incidenza notevolmente maggiore
rispetto alle altre popolazioni mediterranee da loro considerate, ipotizzano
che la rilevante presenza della mutazione
039 nel centro della
Corsica (88,40%) ed in Sardegna (95,40%), incidenza notevolmente maggiore
rispetto alle altre popolazioni mediterranee da loro considerate, ipotizzano
che la rilevante presenza della mutazione ![]() 039
nelle due isole sia una conseguenza dell’azione dell’effetto del fondatore.
039
nelle due isole sia una conseguenza dell’azione dell’effetto del fondatore.
L’azione dell’effetto del fondatore sulla struttura genetica dei Sardi è stata recentemente ripresa anche da Chiarelli (2003), da Francalacci et al. (2003) e da Rootsi et al. (2004). L’effetto fondatore viene ipotizzato da questi Autori in base all’alta incidenza (33-41%) in Sardegna della mutazione M26 del cromosoma Y (Semino et al., 2000; Passarino et al., 2001; Francalacci et al., 2003; Zei et al., 2003; Rootsi et al., 2004). M26 è una mutazione successiva della mutazione M170, entrambe definiscono l’aplogruppo Eu8.
La mutazione M170 avrebbe avuto origine in Europa circa 23000-28000 anni fa (Semino et al., 2000; Chiarelli, 2003; Rootsi, 2004) in popolazioni di cultura Gravettiana, la successiva mutazione M26 avrebbe avuto origine in una popolazione rifugiatasi in Spagna e Francia del Sud durante “L’ultimo Massimo Glaciale” (Last Glacial Maximum: LGM) e che si sarebbe diffusa, dopo LMG, all’incirca tra 13000 e 9000 anni fa in Sardegna (Passarino et al., 2001; Rootsi, 2004). La mutazione M26 è stata sinora rilevata, con basse frequenze, in poche altre popolazioni europee: Castigliani (19%), Baschi francesi (8%), Baschi spagnoli (6%) e con percentuali ancora inferiori in Andalusia, in Italia centrale, Inghilterra, Scozia, Irlanda, etc. (Semino et al., 2000; Bosch et al., 2001; Francalacci et al., 2003; Rootsi et al., 2004).
Da notare che la mutazione M26 è presente in tutta la Sardegna (Passarino et al., 2001; Zei et al., 2003), pur raggiungendo una sua incidenza leggermente superiore nell’area centro-orientale dell’Isola (Zei et al., 2003). Tuttavia M26 mostrerebbe un’elevata eterogeneità tra le diverse aree dell’Isola qualora si considerasse le zone d’origine dei cognomi degli individui campionati. In specifico molti cognomi degli individui portatori di questo aplotipo apparterrebbero originariamente alla zona definita “arcaica” dell’Isola (Zei et al., 2003).
Effetto bottleneck
È ben noto che nel caso in cui una popolazione non sia largamente numerosa o subisca una drastica riduzione le frequenze alleliche possono essere soggette a delle fluttuazioni casuali (genetic drift), che avvengono in misura direttamente proporzionale alla limitatezza della popolazione (Borgognini Tarli e Masali, 1987). La deriva genetica dunque può cambiare il pool genico di una popolazione in base al caso. In conseguenza di una drastica riduzione della popolazione, in genere a seguito di epidemie virulente o per emigrazioni di massa, si assiste ad una forma di deriva genetica detta bottleneck.
Effetti bottleneck che potrebbero essere avvenuti in Sardegna:
- Durante il periodo dell'invasione romana. Nel 174 a.C. per illustrare e perpetuare le sue gesta compiute in Sardegna, Tiberio Sempronio Gracco fece collocare una tavola di bronzo, nel tempio di Roma della dea Mater Matuta, in base al testo della tavola riportato da Livio [XLI, 28] 80.000 Sardi furono uccisi o resi schiavi tra il 177 ed 176 a.C. dall’esercito romano. La riduzione della popolazione complessiva potrebbe essere stata di circa il 27% o di oltre il 50% a seconda della stima della numerosità della popolazione complessiva adottata per il periodo, rispettivamente: 300.000 (Pais, 1923) o 150.000 individui (Meloni, 1975).
- Dal 1320 al 1350. Secondo Day (1987b) la popolazione sarebbe stata rispettivamente pari a 190.000 e 89.000 individui, con un decremento dunque del 53%. Decremento attribuito alle epidemie di peste che colpirono la Sardegna nel 1324 (Fermi, 1934) e nel 1347-1348 (Loddo-Canepa, 1933; Fermi, 1934; Del Panta, 1980; Livi, 1984; Day 1987b).
- Dal Censimento fiscale spagnolo del 1583/1603 a quello del 1627. In base ai dati sui "fuochi" riporatati da Serri (1997a) per il Censimento del 1583 (22.771 fuegos viejos), del 1603 secondo Corridore (1902), e per quello del 1627 (19.623 fuegos nuouos) rileviamo in genere un decremento nel numero dei "fuochi" rispetto ai fuegos viejos del 1583/1603. Tale decremento è attribuibile quasi totalmente ad uno dei due Capi in cui era suddivisa amministrativamente la Sardegna: Capo di Sassari e Capo di Cagliari. In specifico il decremento veniva mostrato dal Capo di Sassari dove si registrava una diminuzione soprattutto nei villaggi dell’interno. Si pensa che l’incongruenza dei risultati tra il Capo di Cagliari e il Capo di Sassari abbia portato il viceré a non accettarli e ad affidare un secondo Censimento all’Inquisizione. Quello che insospettì ancora di più il viceré, fu il fatto che il decremento dei "fuochi" risultava più marcato in zone interne dell’Isola che risultavano di difficile controllo. Il Censimento fatto dall’Inquisizione, pur aumentando complessivamente il numero dei “fuochi” (23.245) confermò un loro effettivo notevole decremento in alcune zone montane tra cui la Encontrada de barbaja Ololay e l’Encontrada de Nuero. Questo dato potrebbe testimoniare un effettivo impoverimento demografico di molti centri montani della zona centrale dell'Isola (Serri, 1997a). Per la Encontrada de barbaja Ololay si registra un crollo demografico di circa il 55%, infatti il numero dei "fuochi" del Censimento del 1583/1603 è pari a 3.555 mentre quello dell'Inquisizione del 1627 risulta di 1.607 (Serri, 1997a). Minore è il decremento mostrato dalla Encontrada de Nuero: 38%, da 1.799 a 1.110 “fuochi” (Serri, 1997a). Dunque osserviamo un probabile effetto bottleneck nella popolazione sarda dell’interno pur in un periodo di incremento, seppure contenuto, del numero dei “fuochi” censiti per l’intera Sardegna. Se consideriamo la perimetrazione in zone storico-geografiche dell’Isola proposta da Ghiani Moi (1964) registriamo un forte decremento dei “fuochi” della Barbagia di Ollolai (-48,85%), mentre più limitati appaiono quelli del Nuorese (-10,44%), del Goceano (-7,72%) e del Monte Acuto (-6,33%) (Tabella 1).
- Dal Censimento fiscale spagnolo del 1627 a quello del 1655. Anatra e Puggioni (1997) propongono per il 1627 ed il 1655 una popolazione pari rispettivamente a 297.424 ed a 220.617 individui, con un decremento dunque pari a circa il 26%. Considerando il numero dei “fuochi”, i maggiori decrementi si registrano (Serri, 1997a) per le zone storico-geografiche (Ghiani Moi, 1964) dell’Anglona (-54,85%), Meilogu (-48,86), Nurra (-46,98%), Coracense (-42,81%), Logudoro (-41,61%), Quirra (-39,19%), Baronia di Siniscola (-36,57%), Monte Acuto (-36,47%), Gallura (-34,18%), Marghine (-31,65%), Nuorese (-28,45%), Sarrabus (-26,51%), Barbagia di Seulo (-25,66%) e Barbagia di Ollolai (-25,68) (Tabella 1). Il decremento è stato attribuito ad una grave carestia avvenuta nel 1644 ed alla peste che imperversò nell’Isola dal 1652 al 1656 (Corridore, 1902).
- Dal Censimento fiscale spagnolo del 1678 a quello del 1688. Anatra e Puggioni (1997) stimano per il 1678 una popolazione composta da 284.947 individui, mentre per il 1688 riportano una popolazione di 229.411; pertanto assisteremo ad un decremento pari al 19%. Se invece seguiamo la quantificazione proposta da Corridore (1902): 299.356 individui per il 1678 e 230.321 per il 1688 si rileva un decremento del 23%. Comunque se prendiamo in considerazione il numero dei “fuochi” censiti per villaggio nel 1678 e consideriamo ciascun “fuoco” come composto da 4 individui (Corridore, 1902) e confrontiamo con la relativa numerosità del Censimento del 1688 (Corridore, 1902) notiamo che diverse zone storico-geografiche dell’Isola (Ghiani Moi, 1964) hanno mostrato dei forti decrementi che ne hanno ridotto drasticamente la popolazione, in particolare: Goceano (-64,65%), Quirra (-51,75%), Barbagia di Ollolai (-51,36%), Baronia di Siniscola (-50,81%), Marghine (-49,86%), Baronia di Orosei (-42,61%), Gallura (-41,22%), Nuorese (-39,75%), Coracense (-39,42%), Gerrei (-36,00%) (Tabella 1). Il decremento sarebbe stato causato da una terribile e tragica carestia avvenuta in Sardegna nel 1680-1681 (Del Panta, 1980).
Da notare che dal Censimento del 1655 al Censimento del 1678 in cui si rileva per l’Isola un incremento demografico pari al 29,16%, essendo la popolazione stimata pari a 220.617 individui per il 1655 ed a 284.947 per il 1678 (Anatra e Puggioni, 1997), si registra, considerando le zone storico-geografiche (Ghiani Moi, 1964), il maggiore decremento per il Goceano (-29,91%) ed altri più contenuti per la Trexenta (-11,10%), la Barbagia di Ollolai (-3,15%) e l’Iglesiente (-2,68%).
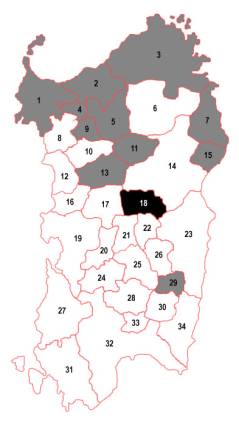
La Barbagia di Ollolai è la zona storico-geografica della Sardegna maggiormente colpita da ripetuti e forti decrementi di popolazione (tra due Censimenti spagnoli almeno del 40%). La Barbagia di Ollolai costituisce il “cuore” della cosiddetta area dei Proto-Sardi in base a quanto riportato da Cappello et al. (1996) (Figura 1).
|
1583-1627 |
1627-1655 |
1678-1688 |
|||
|
Zone storico-geografiche |
|
Zone storico-geografiche |
|
Zone storico-geografiche |
|
|
Barbagia di Ollolai |
-48,85 |
Anglona |
-54,85 |
Goceano |
-64,65 |
|
Nuorese |
-10,44 |
Meilogu |
-48,86 |
Quirra |
-51,75 |
|
Goceano |
-7,72 |
Nurra |
-46,98 |
Barbagia di Ollolai |
-51,36 |
|
Monte Acuto |
-6,33 |
Coracense |
-42,81 |
Baronia di Siniscola |
-50,81 |
|
Sulcis |
-1,85 |
Logudoro |
-41,61 |
Marghine |
-49,86 |
|
Barbagia di Seulo |
0,44 |
Quirra |
-39,19 |
Baronia di Orosei |
-42,61 |
|
Meilogu |
1,23 |
Baronia di Siniscola |
-36,57 |
Gallura |
-41,22 |
|
Barbagia di Belvì |
5,06 |
Monte Acuto |
-36,47 |
Nuorese |
-39,75 |
|
Monte Urtigu |
7,06 |
Gallura |
-34,18 |
Coracense |
-39,42 |
|
Baronia di Orosei |
7,57 |
Marghine |
-31,65 |
Gerrei |
-36,00 |
|
Anglona |
7,74 |
Nuorese |
-28,45 |
Campidano d'Oristano |
-35,22 |
|
Planargia Montiferru |
10,95 |
Sarrabus |
-26,51 |
Anglona |
-33,61 |
|
Coracense |
11,25 |
Barbagia di Seulo |
-25,86 |
Barbagia di Belvì |
-31,10 |
|
Logudoro |
11,37 |
Barbagia di Ollolai |
-25,68 |
Zona Tirso |
-30,16 |
|
Mandrolisai |
11,87 |
Campu Giavesu |
-23,44 |
Parteolla |
-27,15 |
|
Campu Giavesu |
13,33 |
Monte Urtigu |
-21,93 |
Marmilla |
-24,75 |
|
Ogliastra |
14,67 |
Ogliastra |
-19,66 |
Meilogu |
-22,83 |
|
Gallura |
15,65 |
Goceano |
-17,07 |
Planargia Montiferru |
-22,49 |
|
Sarrabus |
16,46 |
Zona Tirso |
-11,94 |
Barbagia di Seulo |
-20,44 |
|
Marmilla |
22,37 |
Sulcis |
-10,15 |
Ogliastra |
-19,02 |
|
Trexenta |
25,60 |
Campidano di Cagliari |
-9,30 |
Arborea |
-18,61 |
|
Sarcidano |
25,62 |
Parteolla |
-8,24 |
Mandrolisai |
-18,48 |
|
Marghine |
28,25 |
Mandrolisai |
-7,96 |
Trexenta |
-17,62 |
|
Campidano d'Oristano |
31,66 |
Sarcidano |
-4,46 |
Logudoro |
-17,43 |
|
Zona Tirso |
33,63 |
Arborea |
-4,08 |
Sarrabus |
-17,36 |
|
Baronia di Siniscola |
34,29 |
Gerrei |
-1,74 |
Campu Giavesu |
-16,75 |
|
Gerrei |
35,90 |
Campidano d'Oristano |
-0,72 |
Sarcidano |
-15,56 |
|
Arborea |
40,65 |
Baronia di Orosei |
3,61 |
Monteleone |
-11,48 |
|
Nurra |
48,87 |
Barbagia di Belvì |
4,57 |
Monte Urtigu |
-9,17 |
|
Iglesiente |
50,66 |
Marmilla |
11,87 |
Nurra |
-8,60 |
|
Campidano di Cagliari |
52,94 |
Planargia Montiferru |
18,69 |
Monte Acuto |
-2,90 |
|
Quirra |
58,57 |
Trexenta |
22,07 |
Sulcis |
-0,81 |
|
Monteleone |
64,74 |
Monteleone |
34,48 |
Iglesiente |
3,75 |
|
Parteolla |
89,58 |
Iglesiente |
67,51 |
Campidano di Cagliari |
3,92 |
Tabella 1. Variazioni percentuali (![]() )
dei “fuochi” nelle zone storico-geografiche della Sardegna tra i Censimenti del
1583/1603, 1627, 1655 e della popolazione tra il Censimento del 1678 e quello
del 1688.
)
dei “fuochi” nelle zone storico-geografiche della Sardegna tra i Censimenti del
1583/1603, 1627, 1655 e della popolazione tra il Censimento del 1678 e quello
del 1688.
Altri decrementi di popolazione, e dunque altri effetti bottleneck nelle comunità sarde, possono essere avvenuti anche in altri periodi (Angioni et al., 1997) e ciò rafforza l’ipotesi che la specificità biologica mostrata dalla popolazione delle zone interne dell’Isola, possa essere una conseguenza dell’azione congiunta della deriva genetica e degli elevati tassi di endogamia e consanguineità registrati per queste comunità sino alla metà (primo quarto) del XX secolo (Sanna et al., 2004a, b).
Flusso genico interno
Un ruolo importante sulla struttura biologica della popolazione sarda potrebbero averlo avuto anche i flussi migratori interni verificatisi nell'Isola nel corso dei millenni. I flussi interni, oltre a quelli probabilmente avvenuti nei periodi: protostorico, punico (535-238 a.C.) e romano (238 a.C. - 455 d.C.), si sarebbero verificati intorno al VII secolo d.C., con un movimento verso le aree interne, nell'XI-XIII secolo verso le pianure, nel XIII secolo verso le città, nel XIV-XV secolo verso le aree interne, tra la fine del XVII e gli inizi del XVIII verso le pianure, agli inizi del XIX verso le città, nella seconda metà del XX secolo verso le città e le zone costiere (Sanna e Melis, 1998; Floris e Sanna, 2000). Da rimarcare che dal 1861 al 1991 le aree di montagna hanno perso circa un 20% della propria popolazione in favore delle zone di pianura (Zei et al., 2003). Inoltre ricordiamo che dal XVII secolo sono state ripopolate da Sardi le seguenti zone storico-geografiche, spopolatate dal XIV al XVI secolo: Nurra, Sarrabus e Sulcis, mentre la Gallura è stata ripopolata al 75% da individui provenienti dalla Corsica meridionale (Terrosu Asole, 1980).
Questi flussi potrebbero aver favorito un processo di omogeneizzazione ridistribuendo i marcatori genetici nello spazio dagli ancestrali luoghi di insediamento.
Figura 1. Zone storico-geografiche della Sardegna (Ghiani Moi, 1966, pp. 35-42) e perimetrazione della cosiddetta “Area arcaica” dell’Isola (Cappello et al., 1986, p. 134).
|

Conclusioni
L’attuale struttura genetica dei Sardi potrebbe essere stata determinata fondamentalmente dall'azione di diversi fattori microevolutivi che hanno agito nel tempo:
1) l’azione dell’effetto del fondatore nella espansione dalla popolazione paleolitica ancestrale e nella fondazione di nuove comunità;
2) l’azione dell’effetto bottleneck in conseguenza di forti decrementi demografici avvenuti nel tempo in Sardegna, particolarmente rilevanti in alcune zone interne dell’Isola;
3) l'isolamento geografico e dunque la relativa scarsità di flussi migratori, in tempi storici, numericamente tali da modificare radicalmente la struttura biologica della popolazione autoctona tramite mescolanza tra le popolazioni o rimpiazzamento degli autoctoni;
4) gli elevati livelli di endogamia e consanguineità, soprattutto delle popolazioni residenti nelle aree montane interne dell’Isola (Sanna et al., 2004a, b), che hanno contribuito ad aumentare l'omozigosi di queste popolazioni.
5) i flussi migratori interni che potrebbero aver determinato un processo di omogeneizzazione, ridistribuendo i marcatori genetici nello spazio dagli ancestrali luoghi di insediamento.
Dunque sembrerebbe potersi affermare che sia la struttura biologica dei Sardi nel loro complesso sia quella della popolazione stanziata nella cosiddetta “Archaic area” dell’Isola potrebbe essere stata condizionata dall’azione congiunta di diversi fattori microevolutivi. Pertanto, per analizzare meglio le differenze interne tra i Sardi e le relative relazioni biologiche con le altre popolazioni sarebbe opportuno che i futuri studi sulla struttura biologica della popolazione sarda siano basati possibilmente su campionamenti ampi e con l’origine degli individui componenti il campione accertata tramite genealogia. Ciò consentirebbe di avere dei dati di base chiaramente riferibili ad un determinato Comune dell’isola al fine di evitare che le induzioni diventino deduzioni.
Acknowlegements. Work supported by MURST 60% grants.
Bibliografia
Anatra, B., e Puggioni, G., 1997 Fonti ecclesiastiche per lo studio della popolazione della Sardegna centro-meridionale. In: Storia della popolazione in Sardegna nell'epoca moderna, a cura di B. Anatra, G. Puggioni, G. Serri, (Cagliari: AM&D) pp. 25-63.
Angioni, D., Loi, S., e Puggioni, G., 1997, La popolazione dei Comuni sardi dal 1688 al 1991 (Cagliari: CUEC).
Angius, A., Melis, P.M., Morelli, L., Petretto, E., Casu, G., Maestrale, G.B., Frumene, C., Bebbere, D., Forabosco, P., e Pirastu, M., 2001, Archival, demographic and genetic studies define a Sardinian sub-isolate as suitable model for mapping complex traits. Human Genetics, 109: 198-209.
Besta, E., 1908-1909, La Sardegna Medioevale. Le istituzioni politiche, economiche, giuridiche, sociali, Vol. II. (Bologna: Forni).
Borgognoni Tarli S., e Masali, M., 1987, Antropologia e Antropometria. (Torino: UTET).
Bosch, E., Calafelli, F., Comas, D., Oefner, P.G., Underhill, P., e Bertranpetit, J., 2001, High-resolution analysis of human Y-chromosome variations shows a sharp discontinuity and limited gene flow between northwestern Afirca and the Iberian peninsula. American Journal of Human Genetics, 68: 1019-1029.
Caocci, A., 1985, La Sardegna (Milano: Mursia Ed.) p. 105.
Cappello, N., Rendine S., Griffo R., Mameli, G.E., Succa, V., e Vona, G., 1996, Genetic analysis of Sardinia: I. Data on 12 polymorphisms in 21 linguistic domains. Annals of Human Genetics, 60: 125-141.
Casula, F.C., 1980, Giudicati e curatorie. In: Atlante della Sardegna, Fasc. II, a cura di R. Pracchi, A. Terrosu Asole, (Roma: Kappa) pp. 94-109.
Cavalli Sforza, L.L., Menozzi, P., e Piazza A., 1994, The history and geography of human genes (Princeton: Princeton University Press).
Chiarelli B., 2003, Dalla natura alla cultura. Principi di Antropologia Biologica e Culturale. Vol. I (Padova: Piccin).
Corridore, F., 1902, Storia documentata della popolazione di Sardegna (1479-1901). (Torino: C. Clausen).
Day, J., 1982, Alle origini della povertà rurale. In: Contadini e pastori nella Sardegna tradizionale, pp. 13-32 (Cinisello Balsamo: CRS-Pizzi).
Day, J., 1987, Uomini e terre nella Sardegna coloniale XII-XVIII secolo (Torino: CELID).
Del Panta, L., 1980, Le epidemie nella storia demografica italiana (secoli XIV - XIX) (Torino: Loescher).
Falchi, A., Giovannoni,
L., Vacca, L., Latini, V., Vona, G., e Varesi, L., 2005, β-globin gene
cluster haplotypes associated with ![]() -thalassemia
on Corsica island. American Journal of Hematology, 78: 27-32.
-thalassemia
on Corsica island. American Journal of Hematology, 78: 27-32.
Falchi, A., Vacca, L., Lopez Alomar, A., Esteban, E., Memmì, M., Varesi, L., Moral, P., e Vona, G., 2004, Population variability in some genes involving the haemostatic system: data on the general population of Corsica (France), Sardinia and Sicily (Italy). Genetics and Molecular Biology, 27: 139-146.
Fermi, C., 1934, Regioni malariche, decadenza risanamento e spesa “Sardegna” (Roma: Tip. Editrice di Roma)
Floris, G., e Sanna, E., 2000, On the biological structure of the Sardinian people. In: Biennal Books of EAA. Vol. 1. Human population genetics in Europe, Susanne C. and Bodzsar E.B. Eds, (Budapest: Eötvös University Press) pp. 85-106.
Francalacci, P., Morelli, L., Underhill, P.A., Lillie, A.S., Passarono, G., Useli, A., Madeddu, R., Paoli, G., Tofanelli, S., Calò, C.M., Ghiani, M.E., Varesi, L., Memmì, M., Vona, G., Lin, A.A., Oefner, A., e. Cavalli Sforza, L.L., 2003, Peopling of three mediterranean islands (Corsica, Sardinia, and Sicily) inferred by Y-chromosome biallelic variability. American Journal of Physical Anthropology, 121: 270-279.
Loddo Canepa, F., 1933, Lo spopolamento della Sardegna durante la dominazione aragonese e spagnola. Atti del Congresso Internazionale per gli Studi sulla popolazione, Vol. I, pp. 651-680 (Roma: Istituto Poligrafico dello Stato).
Ghiani Moi, P., 1964, Sardegna d’oggi (Roma: La Poliedrica)
Ghiani, E., e Vona, G., 2002, Y-Chromosome-specific microsatellite variation in a population sample from Sardinia (Italy). Collegium Antropologicum, 26: 387-401.
Lampis, R., Morelli, L., De Virgilis, S., Congia, M., e Cucca, F., 2000, The distribution of HLA aplotypes reveals that the Sardinian population is genetically differentiated from the other Caucasian populations. Tissue Antigens, 56: 515-521.
Livi, C., 1984, La popolazione della Sardegna nel periodo Aragonese. Archivio Storico Sardo, 34: 24-130.
Latini, V., Vacca, L.,
Ristaldi, M.S., Marongiu, M.F., Memmì, M., Varesi, L., e Vona, G., 2003, ![]() -globin gene cluster haplotypes in the
Corsican and Sardinian populations. Human Biology, 75: 855-871.
-globin gene cluster haplotypes in the
Corsican and Sardinian populations. Human Biology, 75: 855-871.
Meloni, P., 1975, La Sardegna romana (Sassari: Chiarella).
Memmì, M., Moral, P., Calò, C.M., Autori, L., Mamali, G.E., Succa, V., Varesi, L., e Vona, G., 1998, Genetic structure of southwestern Corsica (France). American Journal of Human Biology, 10: 567-577.
Mori, A., 1975, La Sardegna. Le Regioni d'Italia, Vol. XVIII. Seconda edizione. (Torino: Unione Tipografica).
Pais, E., 1923, Storia della Sardegna e della Corsica durante il dominio romano. (Roma: Nardecchia).
Passarino, G., Underhill, P.A., Cavalli Sforza, L.L., Semino, O., Pes, G., Carru, L., Ferrucci, L., Franceschi, C., Deiana, L., Baggio, G., e De Benedictis, G., 2001, Use of Y-chromosome binary markers to study the high prevalence of males in Sardinian centenarians and the genetic structure of the Sardinian population. Human Heredity, 52: 136-139.
Piras I.,
Vona, G., Falchi, A., Latini, V., Ristaldi, S., Vacca, L., Varesi, L., e Calò,
C., 2005, ![]() -globin cluster
haplotypes in normal individuals and
-globin cluster
haplotypes in normal individuals and ![]() °39-thalassemia
carriers from Sardinia, Italy. American Journal of Human Biology, 17: 765-772.
°39-thalassemia
carriers from Sardinia, Italy. American Journal of Human Biology, 17: 765-772.
Quintana-Murci, L., Veitia, R., Fellous, M., Semino, O., e Poloni, E.S., 2003, Genetic structure of Mediterranean populations revelead by Y-chromosome haplotype analysis. American Journal of Physical Anthropology, 121: 157-171.
Rootsi, S., Magri, C., Kivisild, T., Benuzzi, G., Help, H., Bermisheva, M., Kutuev, I., Barac, L., Pericic, M., Balanovsky, O., Pshenicov, A., Dion, D., Grobei, M., Zhivotovsky, L.A., Battaglia, V., Achilli, A., Al-Zahery, N., Parik, J., King, R. Cinnioglu, C., Khusnutdinova, E., Rudan, P., Balanovska, E., Scheffrahn, W., Simonescu, M., Brehm, A., Goncalves, R., Rosa, A., Moisan, J.P., Chaventre, A., Ferak, V., Furedi, S., Oefner, P.J., Shen, P., Beckman, L., Mikerezi, I., Terzic, R., Primorac, D., Cambon-Thomsen, A., Krumina, A., Torroni, A., Underhill, P.A., Santachiara-Benerecetti, A.S., Villems, R., e Semino, O., 2004, Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in Europe. American Journal of Human Genetics, 75: 128-137.
Sanna, E., e Melis, M., 1998, Numerosità della popolazione complessiva in Sardegna dalla Protostoria al Censimento del 1991. In: L’Uomo in Sardegna. Aspetti di Antropobiologia ed Ecologia umana (a cura di G. Floris, E. Sanna) pp. 61-98 (Sestu: Zonza Editori).
Sanna E., Iovine, M.C., Calò, C., e Floris, G., 2004b Evolution of marital structure from 1800 to 1974 in Sardinian altitudinal zones. Biométrie Humaine et Anthropologie, 22: 179-186.
Sanna E., Iovine, M.C., e Floris, G., 2004a Evolution of marital structure in 20 Sardinian villages from 1800 to 1974. Anthropologischer Anzeiger, 62: 169-184.
Semino, O., Passarono, G., Oefner, P.J., Lin, A.A., Arbuzova, S., Beckman, L.E., De Benedictis G., Francalacci, P., Kouvatsi, A., Limborska, S., Marcikiæ, M., Mika, A., Mika, B., Primorac, D., Sanrachiara-Benerecetti, A.S., Cavalli-Sforza, L.L., Underhill, P.A., 2000, The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y-chromosome perspective. Science, 290: 1155-1159.
Serri, G., 1997a Due Censimenti inediti dei “fuochi” sardi: 1583, 1627. In: Storia della popolazione in Sardegna nell'epoca moderna (a cura di B. Anatra, G. Puggioni, G. Serri) pp. 79-112 (Cagliari: AM&D).
Serri, G., 1997b, Il Censimento dei “fuochi” sardi del 1655. In: Storia della popolazione in Sardegna nell'epoca moderna (a cura di B. Anatra, G. Puggioni, G. Serri) pp. 123-144 (Cagliari: AM&D).
Terrosu Asole, A., 1980, Abitati nati o abbandonati tra l'Alto Medioevo e i nostri giorni. In: Atlante della Sardegna (a cura di R. Pracchi, A. Terrosu Asole) pp. 118-136., Fasc. II (Roma: Kappa).
Terrosu Asole, A., 1982, I paesaggi di montagna e il manto boschivo. In: La Sardegna, La geografia, Vol.1, pp. 41-50 (Cagliari: Della Torre).
Varesi, L., Memmì, M., Cristofari, M.C.,. Mameli, G.E., Calò, C.M., e Vona, G., 2000, Mitochondrial control-region sequenze variation in the Corsican population, France. American Journal of Human Biology, 12:339-351.
Vona, G., 1995, Le caratteristiche genetiche dei Sardi. Antropologia Contemporanea, 18:101-109.
Vona, G., Memmì, M., Calò, C.M., Latini, V., Vacca, L., Succa, V., Ghiani, M.E., Moral, P., e Varesi, L., 2002, Genetic structure of the Corsican population: A review. Recent Research Developments in Human Genetics, 1: 147-164.
Vona, G., Moral, P., Memmì, M., Ghiani, G.E., e Varesi, L., 2003, Genetic structure and affinities of the Corsican population (France): classical genetic markers analysis. American Journal of Human Biology, 15: 151-163.
Zei, G., Lisa, A., Fiorani, O., Magri, C., Quintana-Murci, L., Semino, O., Santachiara-Benerecetti, A.S., 2003, From surnames to the history of Y chromosomes: the Sardinian population as a paradigm. European Journal of Human Genetics, 11: 802-807.