
Colloques du Groupement des Anthropologistes de Langue Française (GALF)
Mazières, S., Guitard, E., Sevin, A., Joly, N., Dugoujon, J.M., Salzano, F., Larrouy, G., Crubézy, E., 2006, Structure génétique et histoire biologique de trois populations amérindiennes de Guyane française. Antropo, 11, 51-59. www.didac.ehu.es/antropo
Structure génétique et histoire biologique de trois populations amérindiennes de Guyane française
Genetic structure and biological history of three Amerindian populations of French Guiana
Stéphane Mazières1, Evelyne Guitard1, André Sevin1, Nicolle Joly3, Jean-Michel Dugoujon1, Francisco Salzano2, Georges Larrouy1, Eric Crubézy1
1 Centre d’Anthropologie, UMR 8555 CNRS, Université Paul Sabatier, Toulouse, France.
2 Departamento de Genetica, Instituto de Biociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brazil.
3 Hôpital de Cayenne; Guyane française
Auteur chargé de la correspondance: Pr. Crubézy Eric. Centre d’Anthropologie, Faculté de Médecine. 37, allées Jules Guesde. 31000 Toulouse, France. E-mail: crubezy.eric@free.fr
Mots clés: populations amérindiennes, Guyane française, ADNmt, dérive génétique
Key words: amerindian populations, French Guiana, mtDNA, genetic drift
Résumé
Après avoir extrait l'ADN des sérums de trois populations de Guyane française (Kaliña, Palikur, Emerillon), nous avons sélectionné 102 individus d'après les généalogies établies, et nous nous sommes attachés à reconnaître les principaux haplogroupes mitochondriaux amérindiens. Des trois populations étudiées, seuls les Kaliña possèdaient les quatre lignées mitochondriales amérindiennes. Chez les Palikur, il manquait les haplogroupes A et C et chez les Emerillon, les lignées C et D. L’absence des ces lignées est tout à fait étonnante ; elle pourrait être liée chez les Palikur à leur isolement génétique liée à une structure sociale très endogame tandis que chez les Emerillon le dramatique goulot d'étranglement qui a marqué leur histoire est certainement à prendre en compte. Ces résultats préliminaires nous font penser que le peuplement humain initial du plateau des Guyanes semble présenter face au bassin amazonien, des particularités liées au moins en partie à sa position excentrée comme à certaines de ses structures sociales particulières.
Abstract
DNA from three Amerindian groups living in French Guiana (Palikur Emerillon and Kaliña) has been extracted from sera and a total of 102 individuals selectionned from the genealogical data were examined both by RFLP analysis and by D-Loop sequencing for the mitochondrial Amerindian polymorphisms. The four major founding mtDNA lineages were observed in the Kaliña only. The Palikur lack the A and C natives haplogroups and the C and D lineages were not observed in the Emerillon population. The absence of these lineages is to be noticed. In the Palikur, it could be correlated to their genetic isolation due to their endogam social structure, while in the Emerillon, C and D could have been lost after the drastic bottleneck event they underwent. Those preliminary results suggest that in front of the Amazon bassin, the peopling of the Guianas present pecularities relative to its geographical location as well as some particular social structures.
Introduction
Depuis une quinzaine d'années, les problématiques relatives au peuplement de l’Amérique sont abordées par la biologie moléculaire, notamment par l'étude de l'ADN mitochondrial (ADNmt) (Bonatto et Salzano, 1997; Moraga et al., 2000; Silva Jr., 2002; Salzano, 2002). En effet, son mode de transmission uniparental, son taux de mutation supérieur à l'ADN nucléaire et l'absence de recombinaison génique font de l'ADNmt l'un des outils les plus approprié à l'étude microévolutive des populations (Giles et al., 1980, Anderson et al., 1981). En Amérique, les études ont montré que la quasi-totalité des Amérindiens était regroupée en quatre lignées mitochondriales principales, ou haplogroupes, appelées A, B, C et D (Wallace et al., 1985; Torroni et al., 1992, Horai et al., 1993). Chacun de ces haplogroupes est caractérisé par un polymorphisme de longueur associé à un lot de mutations dans la région hypervariable 1 (HV1) de l'ADNmt. L'haplogroupe A est défini par la présence d’un site de restriction spécifique de l’enzyme HaeIII à la position nucléotidique (np) 663 et les substitutions C16223T, C16390T, G16319A et T16362C. L'haplogroupe B est défini par la délétion de 9 paires de bases dans la partie intergénique COII/tRNALys de la région V de l’ADNmt et les mutations T16189C et T16217C. La lignée C est déterminée par l’absence d’un site HincII au np 13259 et les mutations C16223T, T16298C, T16325C et C16327T. Enfin, les individus de l'haplogroupe D sont caractérisés par l’absence d’un site AluI au np 5176 et les substitutions C16223T, T16325C et T16362C.
L’histoire des populations de Guyane française est relativement peu connue en raison du milieu équatorial qui se prête mal aux prospections archéologiques ainsi qu’à la conservation du matériel. En effet, l’essentiel des données archéologiques relatives à l'occupation précolombienne en Guyane a principalement été retrouvé sur la frange littorale (Rostain, 1992; 1994). Toutefois, suite aux travaux de Larrouy et al. (1964a,b), Daveau et al. (1975), Tchen et al. (1978a,b; 1981) et Dugoujon et al. (1994) menés sur les marqueurs classiques du sang, les premières relations génétiques entre les populations amérindiennes de Guyane Française, y compris celles de l’intérieur inaccessibles par l’archéologie, étaient proposées.
Actuellement, aucune étude au niveau moléculaire n’a été réalisée sur les groupes amérindiens de Guyane. Dans ce travail, nous nous proposons donc d'analyser trois populations Amérindiennes de Guyane française pour les marqueurs de l’ADNmt qui décrivent l'ensemble de la variabilité amérindienne. La confrontation de nos résultats aux données issues de l'ethnologie et de la démographie devrait nous aider à interpréter la composition génétique observée et apporter un éclairage nouveau à la compréhension à l'histoire du peuplement de la Guyane.
La Guyane et ses populations amérindiennes
La Guyane française est caractérisée par un fort contraste géographique et démographique. Le littoral, où 90% de la population vit, est constitué de bancs sableux flanqués par les boues charriées par l'Amazone. Le reste du département, recouvert par la forêt sempervirente, est peuplé de manière éparse. Cinq groupes amérindiens vivent actuellement en Guyane française, deux sur le littoral et trois dans la partie intérieure du département (figure 1).
Les populations de l’intérieur
Les Wayãpi, Wayana et Emerillon sont les trois groupes indiens de l’intérieur de la Guyane. Leur culture et leur mode de vie sont adaptés à la forêt tropicale, avec notamment la pratique de l’agriculture sur brûlis, de la chasse, de la pêche et de la cueillette (Grenand et Grenand 1978, 1985b,c,d; Grenand, 1981).
Wayãpi
Ces Indiens de langue Tupi-Guarani seraient venus du bas Amazone et de la région du Xingu pour s'installer en Guyane à la fin du 18ème siècle (Grenand et Grenand, 1985d). Environ 500 personnes vivent réparties en deux communautés le long de la rivière Oyapock. Deux cent dix-huit vivent dans deux villages de la région de Camopi et deux cent quatre vingt trois habitent trois villages près de Trois-Sauts. Le choix du conjoint se fait entre cousins croisés ou entre cousins croisés classificatoires. Le taux d’endogamie y est assez élevé et le métissage avec des groupes non Indiens est peu fréquent (Dugoujon et al., 1994).
Wayana
Ce groupe Indien de langue Karib, complètement isolé jusqu’au milieu du 19ème siècle, a été moins affecté par les épidémies. Par conséquent, les Wayana sont les plus nombreux des Indiens de l’intérieur (Grenand et Grenand, 1985c). Environ 550 personnes vivent dispersées dans 10 villages le long du haut Maroni et de la rivière Litani. Le système matrimonial des Wayana repose sur le choix préférentiel de cousins croisés classificatoires, ce qui augmente le taux de consanguinité (Bonnet, 1990 ; Dugoujon et al., 1994). Ce système est cependant contre-balancé par la recherche des gendres dans les villages voisins.
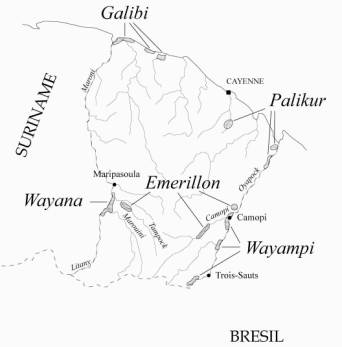
Figure 1. Localisation géographique des populations Amérindiennes de Guyane française.
Figure 1. Geographic location of the French Guiana Natives
Emerillon
Les Emerillon résidaient en Guyane dès le 16ème siècle. Ce groupe de langue Tupi échappe de justesse à l’extinction dans les années 1950 quand leur effectif est réduit à 52 individus (Grenand and Grenand, 1979). Un peu plus de 124 Emerillon occupent aujourd’hui la région de Camopi et les rives du Tampock. (Bonnet, 1990; Dugoujon et al., 1994).
Les populations du littoral
Deux groupes amérindiens occupent le littoral guyanais : les Palikur et les Kaliña. Le remodelage du littoral par les courants marins les contraint à un semi-nomadisme. Ces populations se sont nettement adaptées à leur environnement puisqu’ils maîtrisent depuis longtemps la navigation, particulièrement les Kaliña. Le métissage est rare mais plus fréquent que chez les populations de l’intérieur (Larrouy et al., 1964a).
Palikur
Les Palikur sont la tribu la plus ancienne du littoral. Apparentés au groupe linguistique Arawak, ils seraient originaires du haut Xingu, dans le Sud-ouest de l’Etat de Pará (Brésil). Vincente Pinzon les a d’abord identifiés en 1550 à l’ouest de l’embouchure de l’Amazone. Au 19ème siècle, Henri Coudreau note leur présence sur la rivière Arukawa (Etat de Pará, Brésil) (Arnaud, 1980). Actuellement, 400 sujets se trouvent au bord de l’estuaire de l’Oyapock (la Savane et Trois Palétuviers) et 600 individus vivent dans la vallée de l’Arukawa (Brésil).
Kaliña
Les Kaliña formaient la plus importante tribu Karib de la côte guyanaise, avec 2000 sujets en 1652. Avec l’arrivée des Européens, ils sont employés comme chasseurs d’esclaves. Mais la dysenterie et les maladies pulmonaires ne les épargnent pas. Décimés, il ne reste que 250 sujets en 1848. Les Kaliña survivants s’installent sur la rive gauche du Maroni en Guyane néerlandaise, l’actuel Surinam. Ce n’est qu’en 1780 qu’ils reviennent en Guyane française, où ils sont estimés à 1700 individus en 1985. Les Kaliña occupent actuellement le littoral nord-ouest de la Guyane, au bord de l’estuaire du Maroni et de la Mana (Bonnet, 1990). De tempérament combatif, ils ont longtemps été en conflit avec les tribus voisines, Arawak notamment.
Les communautés Kaliña sont organisées autour d’une ou plusieurs unités patriarcales. Le choix du conjoint se faisait entre cousins croisés. Aujourd’hui, seul le mariage entre cousins croisés classificatoires est toléré ; devenu rare, il laisse place au libre choix (Grenand et Grenand, 1978). Les précédentes investigations ont montré que les Kaliña ont le plus important taux de métissage non-Indien (Larrouy et al., 1964a).
Démographie amérindienne
Les plus anciens documents relatifs aux Indiens de Guyane française remontent au 16ème siècle et la plupart proviennent de missions jésuites, mais les données ethnologiques et démographiques antérieures à 1947 sont généralement imprécises. Les données récentes et fiables dérivent donc essentiellement des travaux de Hurault (1965), Bois (1967), et Grenand et Grenand (1978; 1979; 1985a,b,c,d). Au 17ème siècle, près de 9500 Amérindiens occupaient le littoral guyanais, mais cette population sera réduite à moins de 500 individus dans la deuxième moitié du 18ème siècle. De la même manière, à l'intérieur du département, 300 Wayana et 400 Emerillon ont été recensés en 1770 et 6000 Wayampi en 1824. Ces trois populations verront leur effectif diminuer plus tardivement ; au milieu du 20ème siècle, il ne reste que 900 sujets dans la partie intérieure de la Guyane. Depuis, la démographie amérindienne est en croissance avec environ 3300 Indiens dénombrés à la fin des années 1970 (Grenand and Grenand, 1979).
Matériel et Méthodes
Collecte des échantillons
Entre 1964 et 1985, les Amérindiens de Guyane ont été le centre d’intérêt de missions de terrain menées par le Pr. Larrouy sous les auspices du CNRS (Centre d’Hémotypologie, UMR 8555, Toulouse) avec le soutien de l’Institut Pasteur. Au cours de celles-ci, des prélèvements sanguins ont été réalisés conjointement à des enquêtes généalogiques. Tous les échantillons ont été collectés dans des tubes sur anticoagulants EDTA ou ACD. Les Emerillon ont été prélevés en 1988 par le Dr. Etienne Bois.
Extraction et méthode d’amplification de l’ADN
L'ADN des populations Palikur, Kaliña et Emerillon a été extrait à l’aide du kit NucleoSpin® Blood QuickPure (Macherey-Nagel) à partir des prélèvements sériques collectés sur le terrain et actuellement conservés au Centre d’Hémotypologie de Toulouse (UMR 8555). Chacun des individus a été analysé par PCR-RFLP et séquençage automatique pour les polymorphismes de l'ADN mitochondrial qui décrivent la variabilité génétique amérindienne. Après amplification, le polymorphisme 9pb-deletion, caractéristique des individus B, a été directement lu sur un gel d’agarose à 3%. Les haplogroupes A, C et D ont, quant à eux, été déterminé après digestion enzymatique dans une solution contenant 2U de l’endonucléase appropriée.
Analyse statistique
Les données généalogiques ont permis de tracer les liens de
parenté entre les sujets prélevés et de mettre en évidence 102 lignées
maternelles indépendantes parmi les trois populations étudiées. Les fréquences
des haplogroupes ont été obtenues par comptage à partir de ces lignées. Le taux
d’hétérozygotie a été calculé d’après la formule ![]() .
.
Résultats
Le tableau 1 présente la répartition des haplogroupes amérindiens fondateurs au sein des trois groupes étudiés, ainsi que le taux d’hétérozygotie. On observe clairement une inégalité de la distribution des lignées mitochondriales, puisque les Kaliña possèdent les quatre haplogroupes alors que les lignées C et D, et A et C n’ont pas été retrouvées chez respectivement les Emerillon et les Palikur. Chez ces dernières, l’absence de la moitié de la diversité génétique a un effet direct sur le taux d’hétérozygotie, puisque les Palikur et les Emerillon ont les taux les plus faibles (0,487 et 0,336). Par ailleurs, nous avons observé des individus Kaliña qu’il nous a été impossible de classer parmi les haplogroupes A, B, C et D. Ces individus, désignés comme "autres", représentent près de 7% des lignées mitochondriales Kaliña.
|
Populations |
Haplogroupe |
Hétérozygotie |
||||
|
A |
B |
C |
D |
autre |
H |
|
|
Kaliña |
6,9% |
41,4% |
37,9% |
6,9% |
6,9% |
0,671 |
|
Emerillon |
21,4% |
78,6% |
0 |
0 |
0 |
0,336 |
|
Palikur |
0 |
58,1% |
0 |
41,9% |
0 |
0,487 |
Tableau 1. Distribution des lignées mitochondriales fondatrices chez trois populations de Guyane française.
Table 1. Distribution of the mtDNA founding lineage in three French Guiana Natives.
Discussion
L’étude de trois populations de Guyane française pour les polymorphismes de l’ADNmt qui décrivent l’ensemble de la diversité amérindienne a révélé des particularités génétiques pour chacune d’entre elles. Tout d’abord, la présence de lignées sortant de la variabilité amérindienne chez un seul des groupes étudiés, les Kaliña, amène à se poser la question au sujet de leur provenance. D'après les travaux menés sur près de vingt populations amazoniennes examinées pour les marqueurs classiques du sang, les Kaliña possèdent les fréquences les plus élevées de composantes biologiques exclusivement observées dans l’Ancien Monde (Larrouy et al., 1964a; Black et al., 1988; Callegari-Jacques et al., 1994; Santos et al, 1998; Battilina et al., 2002). Ces allèles ont donc probablement été introduits par métissage, d’autant plus que que les Kaliña vivent à proximité de groupes Créoles et Noirs Marrons (Larrouy et al., 1964a). De la même manière, nous pouvons penser que les individus Kaliña non classés parmi les haplogroupes A, B, C et D ont une ascendance maternelle non-Amérindienne.
Les Palikur et les Emerillon ont ensuite été caractérisés par l’absence de deux lignées fondatrices. La disparition de deux lignées sur quatre dans une même population amérindienne est peu courante puisque seuls les groupes du sud de la Patagonie et de la Terre de Feu (Tehuelche, Selk'nam, Yamana, Kaweshar et Yaghan), représentés par les lignées C et D, font pour cela, figure de référence sur l’ensemble du continent (Lalueza et al., 1997; Moraga et al., 2000; García-Bour et al., 2004). Partant du principe qu’elles possèdent en commun la même caractéristique (absence de A et B), Lalueza et al (1997) et Moraga et al (2000) supposent que ces populations sont étroitement liées et qu’elles proviendraient toutes de la même dynamique de peuplement au cours de laquelle les lignées fondatrices A et B auraient été absentes ou perdues. Le phénomène est toutefois diffèrent chez les Palikur et les Emerillon puisque ce ne sont pas les mêmes haplogroupes qui les représentent. Les lignées A et B ont été observées chez les Emerillon alors que les Palikur ont été définis par la présence de B et D. Contrairement à leurs consœurs du sud, les Palikur et les Emerillon actuels n’ont donc probablement pas suivi la même dynamique de mise en place.
Les données collectées par Grenand et Grenand (1979) sur la démographie des groupes amérindiens de Guyane relatent une diminution brutale et dramatique des effectifs, principalement causée par les épidémies importées de l'Ancien Monde. Mais ces données révèlent surtout que les Emerillon ont subi la plus importante des réductions d’effectifs, puisqu’au milieu du 20ème siècle il ne subsiste que 52 individus. Or, plus l’effectif survivant est petit et moins grande est la probabilité qu’il soit représentatif du groupe de départ en terme de composition génique. Nous pouvons donc imaginer que les haplogroupes C et D, actuellement absents chez les Emerillon, ont été perdus dans ce goulot d’étranglement.
Parallèlement à l'absence des lignées A et C, les Palikur présente une autre caractéristique. Grenand et Grenand (1987) ont en effet décrit une organisation sociale clanique des Palikur, régie par des règles matrimoniales précises de type exoclanique et endogame que les généalogies sur près de 6 générations établies sur le terrain ont faites apparaître. Dans ce type de structure, un individu Palikur ne peut se marier avec un membre de son propre clan puisque ce type d’union est considéré comme incestueux. Le choix du conjoint se fait donc à l’extérieur du clan, mais à la seconde condition que ce conjoint soit lui-même Palikur. Puisque les mariages Palikur unissent exclusivement des individus eux-mêmes Palikur, les échanges avec les génomes des groupes alentours sont donc inexistants. Par conséquent, la composition génétique actuelle des Palikur, avec l’absence des lignées A et C, résulte très probablement de cette barrière et cet isolement génétiques imposés par leurs règles sociales.
Les Palikur, les Emerillon et les Kaliña ont déjà été examinés pour les marqueurs sanguins (Larrouy et al., 1964a,b ; Daveau et al., 1975 ; Potel, 1988), et les résultats ont été comparés à ceux des populations Wayampi et Wayana de Guyane (Tchen et al. ; 1978a,b; 1981 ; Dugoujon et al., 1994) ainsi qu’aux Apalaï-Wayana (Salzano et al., 1988) ; un groupe Amérindien vivant à proximité de la frontière avec la Guyane. La figure 2 représente les relations génétiques obtenues (Mazières et al., 2005 ; soumis pour publication). Ce dendrogramme montre tout d’abord des similitudes génétiques entre les Kaliña, les Wayana, les Wayampi et les Apalaï-Wayana, regroupés dans un cluster central. Mais surtout, il révèle que pour les groupes sanguins, les Palikur et les Emerillon se distinguent fortement des autres populations, ce qui, de manière intéressante, confirme les observations précédemment faites pour l’étude de l’ADNmt quant à la singularité génétique de ces deux populations.
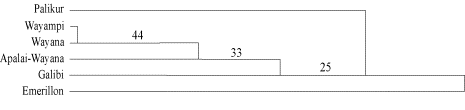
Figure 2. Dendrogramme basé sur douze systèmes sanguins montrant les relations génétiques entre les populations de Guyane et les Apalai-Wayana. Les nombres indiquent les valeurs du bootstrap en pourcentage, basé sur 2000 réplications.
Figure 2. Dendrogram based on twelve blood groups showing the relationships among the French Guiana populations and the Apalai-Wayana. The numbers indicate bootstrap values in percentage, based on 2.000 replications.
Conclusion
Trois populations amérindiennes de Guyane Française ont été analysées pour les polymorphismes de l’ADNmt définissant la diversité génétique amérindienne. La distribution des haplogroupes fondateurs a révélé l’absence des lignées fondatrices A et C chez les Palikur, et C et D chez les Emerillon. Ces particularités appuient les résultats obtenus pour ces mêmes populations par l’étude des groupes sanguins. De plus, elles semblent parfaitement illustrer des évènements ethnohistoriques propres à la Guyane française.
Bibliographie
Anderson, S., Bankier, A.T., Barrell, B.G., de Bruij, M.H.L., Coulson, A.R., Drouin, J., Eperon, I .C., et al., 1981, Sequence and organization of the human mitochondrial genome. Nature, 290, 457-465.
Arnaud, E., 1980, O protestantismo entre os ìndios Palikur do Rio Urucauà (Oiapoque, Brazil) – Notìcia preliminar. Revista de Antropologia (São Paulo), 23, 99-102.
Battilana, J., Bonatto, S.L., Freitas, L.B., Hutz, M.H., Weimer, T.A., Callegari-Jacques, S.M., Batzer, M.A., Hill, K., Hurtado, A.M., Tsuneto, L.T., Petzl-Erler, M.L., Salzano, F.M, 2002. Alu insertions versus blood group plus protein genetic variability in four Amerindian populations, Ann Hum Biol, 29(3), 334-47.
Black, F.L., Santos, S.E.B., Salzano, F.M., Callegari-Jacques, S.M., Weimer, T.A., Franco, M.H.L.P., Hutz, M.H., Rieger, T.T., Kubo, R.R., Mestriner, M.A., Pandey, J.P., 1988, Genetic variation within the linguistic Tupi group: new data on three Amazonian tribes. Ann Hum Biol, 15, 337-351.
Bois, E., 1967. Les Amérindiens de la Haute-Guyane française. Anthropologie, pathologie, biologie (Paris : Editions du C.N.R.S., Desclée Ed.).
Bonatto, S.L. and Salzano, F.M., 1997, A single and early migration for the peopling of the Americas supported by mitochondrial DNA sequence data. Proc. Natl. Acad. Sci. USA, 94,1866-1871.
Bonnet, F., 1990, Analyse des données génétiques relatives à différentes ethnies de la Guyane française – Bilan et interprétation - . Mémoire de DEA d’Anthropologie, option Anthropologie biologique, Université Paul Sabatier, Toulouse III.
Callegari-Jacques, S.M., Salzano, F.M., Weimer, T.A., Hutz, M.H., Black, F.L., Santos, S.E.B., Franco, M.H.L.P., Guerreiro, J.F., Mestriner, M.A., Pandey, J.P., 1994, Further blood genetic studies on Amazonian diversity –data from four Indian groups. Ann Hum Biol 21(5), 465-481.
Daveau, M., Rivat, L., Langaney, A., Afifi, N., Bois, E., Ropartz, C., 1975, Gm and Inv allotypes in French Guiana Indians. Hum Hered.,25(2), 88-92.
Dugoujon, J.M., Guitard, E., Senegas, M.T., Grenand, P., Bois, E., 1994, Gm and Km allotypes in Wayampi, Wayana and Emerillon Indians from French Guiana. Ann. Hum. Biol., 21(4), 335-45.
Garcia-Bour, J., Perez-Perez, A., Alvarez, S., Fernandez, E., Lopez-Parra, A.M., Arroyo-Pardo, E., Turbon, D., 2004, Early population differentiation in extinct aborigines from Tierra del Fuego-Patagonia: ancient mtDNA sequences and Y-chromosome STR characterization. Am J Phys Anthropol, 123(4), 361-70.
Giles, R.E., Blanc, H., Cann, H.M., Wallace, D.C., 1980, Maternal inheritance of human mitochondrial DNA. Proc Natl Acad Sci U S A, 77(11) ,6715-9.
Grenand, F., Grenand, P., 1978, Les populations amérindiennes de Guyane Française. ORSTOM, Cayenne (GUF), 14 p.
Grenand, F., Grenand, P., 1979, Les Amérindiens de Guyane française aujourd’hui : éléments de compréhension. Bull. Soc. Americanistes, LXVI, 361-382.
Grenand, P., 1981, Agriculture sur brûlis et changements culturels : le cas des Indiens Wayapi et Palikur de Guyane. Journal d'Agriculture Traditionnelle et de Botanique Appliquée, 28 (1), 23-31.
Grenand, P., Grenand, F., 1985a, La question amérindienne en Guyane française : Eléments d'histoire amérindienne. Ethnies (FRA), 1(1-2), 11-14.
Grenand, P., Grenand, F., 1985b, La question amérindienne en Guyane française : Les Arawak. Ethnies (FRA), 1(1-2), 15-17.
Grenand, P., Grenand, F., 1985c. La question amérindienne en Guyane française : Les Wayana. Ethnies (FRA). 1(1-2), 23-24.
Grenand, P., Grenand, F., 1985d. La question amérindienne en Guyane française : Les Wayapi. Ethnies (FRA). 1(1-2), 25-26.
Grenand, F., Grenand, P., 1987. La côte d'Amapá, de la bouche de l'Amazone à la baie d'Oyapock, à travers la tradition orale palikur. Boletim do Museu Paraense Emílio Goeldi, série Antropologia, Belém, Pará, 3- 1, 1-78.
Horai, S., Kondo, R., Nakagawa-Hattori, Y., Hayashi, S., Sonoda, S., Tajima, K., 1993, Peopling of the Americas, founded by four major lineages of mtDNA. Mol Biol Evol, 10, 23-47.
Hurault, J., 1965, La population des Indiens de Guyane française. Population, 20, 603-632, 801-828.
Lalueza, C., Perez-Perez, A., Prats, E., Cornudella, L., Turbon, D., 1997, Lack of founding Amerindian mitochondrial DNA lineages in extinct aborigines from Tierra del Fuego-Patagonia. Hum Mol Genet, 6(1), 41-6.
Larrouy, G., Marty, Y., Ruffié, J., 1964a, Etude hémotypologique des populations indiennes de Guyane française. Les groupes érythrocytaires. Bull. Mem. Soc. Anthrop. Paris 71(1), 107-117.
Larrouy, G., Marty, Y., Ruffié, J., 1964b, Etude hémotypologique des populations indiennes de Guyane française. Les groupes sériques du système Gm. Bull. Mem. Soc. Anthrop. Paris 71(1), 119-123.
Mazières, S., Sevin, A., Bonnet, F., Crubézy, E., Dugoujon, J.-M., Salzano, F. M., Larrouy, G., 2005, Genetic studies in French Guiana populations : synthesis. Am J Phys Anthropol (soumis pour publication).
Moraga, M.L., Rocco, P., Miquel, J.F., Nervi, F., Llop, E., Chakraborty, R., Rothhammer, F., Carvallo, P., 2000, Mitochondrial DNA polymorphisms in Chilean aboriginal populations: implications for the peopling of the southern cone of the continent. Am J Phys Anthropol, 113(1), 19-29.
Potel, J.F., 1988, Les Palikur: Structure sociale et hémotypologique. Positionnement génétique parmi les Amérindiens. Mémoire de DEA d’Anthropologie – Université Paul Sabatier, Toulouse III.
Rostain, S., 1992, L’occupation amérindienne ancienne du littoral de Guyane. Dans Evolution des littoraux de Guyane et de la zone caraïbe méridionale pendant le quaternaire, édité par M. T. Prost (ORSTOM, Paris), p. 481-506.
Rostain, S., 1994, L’occupation amérindienne ancienne du littoral de Guyane. Thèse, Université de Paris 1, Paris (FRA), 953 p ; 2 tomes.
Salzano, F.M., Black, F.L., Callegari-Jacques, S.M., Santos, S.E.B., Weimer, T.A., Mestriner, M.A., Pandey, J.P., Hutz, M.H., Rieger, T.T., 1988, Genetic variation within a linguistic group: Apalai-Wayana and other Carib tribes. Am J Phys Anthropol, 75, 347-356.
Salzano, F.M., 2002, Molecular variability in Amerindians: widespread but uneven information. An Acad Bras Cienc, 74(2), 223-63.
Santos, S.E.B., Ribeiro-Dos Santos, A.K.C., Gueerreiro, J.F., Santos, E.J.M., Weimer, T.A., Callegari-Jacques, S.M., Mestriner, M.A., Franco, M.H.L.P., Hutz, M.H., Salzano, F.M., 1998, New proteic genetic studies in six Amazonian Indian populations. Ann Hum Biol, 25 (6), 505-522.
Silva, W.A. Jr., Bonatto, S.L., Holanda, A.J., Ribeiro-Dos-Santos, A.K., Paixao, B.M., Goldman, G.H., Abe-Sandes, K., Rodriguez-Delfin, L., Barbosa, M., Paco-Larson, M.L., Petzl-Erler, M.L., Valente, V., Santos, S.E., Zago, M.A., 2002, Mitochondrial genome diversity of Native Americans supports a single early entry of founder populations into America. Am J Hum Genet, 71(1), 187-92.
Tchen, P., Bois, E., Seger, J., Grenand, P., Feingold, N., Feingold, J.A., 1978a, Genetic study of two French Guiana Amerindian populations. I. Serum proteins and red cell enzymes. Hum Genet, 45(3), 305-15.
Tchen, P., Seger, J., Bois, E., Grenand, P., Fribourg-Blanc, A. and Feingold, N., 1978b, Genetic study of two French Guiana Amerindian populations.II. Rare electrophoretic variants. Hum Genet, 45, 317-326.
Tchen, P., Bois, E., Lanset, S., Feingold, N., 1981, Blood group antigens in the Emerillon, Wayampi, and Wayana Amerindians of French Guiana. Hum Hered, 31(1), 47-53.
Torroni, A., Schurr, T.G., Yang, C.-C., Szathmary, E.J.E., Williams, R.C., Schanfields, M.S., Troup, G.A., Knowler, W.C., Lawrence, D.N., Weiss, K.M. and Wallace, D.C., 1992, Native American Mitochondrial DNA Analysis Indicates That the Amerind and the Na-Dene Populations Were Founded by Two Independant Migrations. Genetics, 130, 153-162.
Wallace, D.C., Garrison, K., Knowler, W.C., 1985, Dramatic founder effects in Amerindian mitochondrial DNAs. Am J Phys Anthropol, 68(2), 149-55.