
Colloques du Groupement des Anthropologistes de Langue Française (GALF)
Guipert, G., Mafart, B., 2006, Analyse morphométrique d’un os isolé et paléoanthropologie: intérêts et limites, 11, 285-291. www.didac.ehu.es/antropo
Analyse morphométrique d’un os isolé et paléoanthropologie: intérêts et limites
Morphometric studies of an isolated bone in paleoanthropology: interest and limits
Gaspard Guipert et Bertrand Mafart
Antenne du Département de Préhistoire du Muséum National d’Histoire Naturelle, Géosciences de l’Environnement, CEREGE, Europole Méditerranéen de l’Arbois, Bâtiment Villemin, 13545, Aix en Provence, Cedex 04.
Auteur chargé de la correspondance: Gaspard Guipert, Antenne du Département de Préhistoire du Muséum National d’Histoire Naturelle, Géosciences de l’Environnement, CEREGE, Europole Méditerranéen de l’Arbois, Bâtiment Villemin, 13545, Aix en Provence, Cedex 04. Email: gaspard_guipert@hotmail.com
Mots clés: Modélisation tridimensionnelle; Superimposition Procuste; Os pariétal; Voûte bipariétale.
Key words: 3D Modelisation; Procuste Superimposition; Parietal bone; Biparietal vault.
Résumé
Le développement des techniques de comparaisons Procuste a entraîné en paléoanthropologie la multiplication des analyses morphométriques, surtout 2D. Nous avons considéré un os isolé (pariétal) dans un cadre tridimensionnel et étudié une série d’Hominidés fossiles allant des Australopithecus africanus aux Homo sapiens. Nous avons aussi procédé à la modélisation virtuelle de la voûte bipariétale. Les résultats des analyses en composantes principales réalisées après superimpositions Procuste dans les deux cas (modélisation os isolé puis bipariétal), permettent de mettre en évidence: a/ une possibilité de discrimination de certains phylums à partir de la seule analyse de l’os pariétal isolé, b/ mais aussi une nécessité de multiplier les modélisations pour améliorer les discriminations entre ceux-ci.
Abstract
The development of the Procuste techniques in paleoanthropology involved a multiplication of morphometric studies, especially 2D. Here, we considered an isolated bone (the parietal bone) within a 3D framework and studied it on Hominid fossils, from Australopithecus africanus to Homo sapiens. We also proceeded to virtually model the biparietal vault. For both the isolated bone and biparietal vault, the results of the PCA carried out after Procuste superimpositions highlight: a/ a possibility to discriminate certain phyla from the analysis of the isolated parietal bone alone, b/ but also a need for multiplying modelings to improve phyletic discriminations.
Le développement des outils d’investigation numériques (scanner médical, digitaliseur portable) a généré un nouvel axe de recherche pour la paléoanthropologie: l’analyse morphométrique tridimensionnelle au moyen des techniques Procuste (Bookstein, 1989, 1991; Penin et Baylac, 1995; Bookstein, 1996, 1997; Mafart, 2002; Zollikofer, 2002; Zollikofer et al., 2002; Bookstein et al., 2003; Mafart et al., 2004). Parmi les nombreux travaux issus de ce développement (Bookstein et al., 1999; Delson et al., 2001) un certain nombre se focalise plus sur la modélisation tridimensionnelle et l’étude d’ossements isolés (Harvati, 2002, 2003; Guipert et Mafart, sous presse). La localisation tridimensionnelle de points de repères ou landmarks permet de comparer différentes populations fossiles ou du moins les caractères de cette population au moyen d’analyses en composantes principales (ACP) ou autres analyses multivariées.
De part sa robustesse, le pariétal est un os souvent retrouvé lors des fouilles, soit en articulation, soit isolé. Lorsque l’on retrouve d’autres fragments crâniens du même individu, des difficultés d’assemblage peuvent apparaître, liées aux différentes angulations entre les pièces osseuses (zone d’articulation limitée). L’imagerie 3D permet d’obtenir virtuellement des symétriques des pariétaux conservés, de mettre en connexion les images tomodensitométriques 3D de ces pariétaux avec leur symétriques et donc de reconstituer les voûtes bipariétales. Le but de cette étude était d’évaluer les possibilités de discrimination phylétiques comparées des pariétaux et des voûtes bipariétales.
Matériel
Nous avons réalisé des modélisations virtuelles à partir d’images scanographiques de la face externe d’os pariétaux isolés ou prélevées virtuellement sur des fossiles (originaux et moulages, collection du laboratoire d’Anthropologie de la Faculté de Médecine, secteur Nord, Marseille; figure 1).
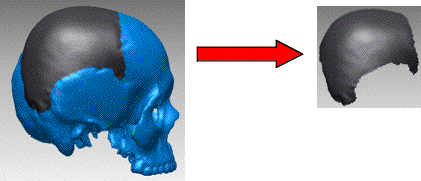
Figure 1: Prélèvement virtuel de l’os pariétal droit sur une modélisation de crâne d’Homo sapiens.
L’étude a porté sur 157 calottes crâniennes (moulages et originaux) et 15 pariétaux droits d’Hominidés fossiles (dont Arago 47) et modernes (population historique de Notre-Dame-du-Bourg, Dignes, France, collection du laboratoire d’Anthropologie de la Faculté de Médecine, secteur Nord, Marseille; effectifs utilisés en tableau 1).
|
Pariétaux: Espèces |
Isolés |
Prélevés virtuellement |
|
Australopithèques (Sts 5, Sts 71) |
0 |
2 |
|
Early Homo (KNM-ER 1470, KNM-ER 1813, OH 7) |
1 |
2 |
|
H. ergaster et H. erectus africain (KNM-ER 3733, KNM-ER 3883, KNM-WT 15000, OH 9) |
1 |
3 |
|
H. erectus asiatiques (Dali, Hexian, Narmada, Ngawi 1, Sambungmacan 1, 3, Sangiran 2, 3, 10, 17, 38, Ngandong 5, 6, 7, 9, Trinil 2, Zhoukoudian III, XI, XII) |
2 |
17 |
|
H. cepranensis (Ceprano 1) |
0 |
1 |
|
H. heidelbergensis d’Europe et d’Afrique (Arago 47, BSV 2, Kabwe, Elandsfontein, Petralona, Sima de los Huesos 5, Swanscombe, Ternifine 4) |
1 |
7 |
|
H. neanderthalensis (Amud 1, Cova Negra, Engis 2, La Chapelle-aux-Saints, La Ferrassie 1, La Quina H5, La Quina H18, Monte Circeo 1, Neanderthal 1, Saccopastore 1, Teshik Tash) |
1 |
10 |
|
H. sapiens préhistoriques (Chancelade 1, Cro-Magnon 1, 2, 3, Hotu Cave 2, Jebel Irhoud 1, LH 18, Markina Gora, Mladec V, Obercassel 2, Pataud, Predmost III, Qafzeh 9, Skhul 5) |
0 |
14 |
|
Hommes modernes |
9 |
100 |
Tableau 1: Effectifs étudiés (originaux soulignés)
Méthode
Les données surfaciques et volumiques ont été obtenues au moyen de scanners médicaux (format DICOM sur un appareil Général Electrique LightSpeed 16, service Scanographie du Professeur Agostini, Hôpital Sainte Marguerite, Marseille). Nous avons réalisé une hélice en coupes de 1,25 mm reconstruite tous les 0,625 mm avec un temps de rotation de 0,7 tour/s, une matrice 512x512 pixels, un SFOV (Field of view) de 25cm, soit une résolution de 0,48 mm. Les ossements ont été modélisés avec le logiciel Mimics ® et des logiciels développés pour le projet FOVEA (http://foveaproject.free.fr) sous la direction de B. Mafart.
Les points de repères craniométriques (Martin et Saller, 1957; Delattre et Fénart, 1960; Grimaud-Hervé et Jacob, 1983; Guipert, 2005) ont été coordonnés tridimensionnellement, soit directement sur les reconstructions virtuelles, soit au moyen d’un digitaliseur portable, Microscribe3DX ®, puis ont été ensuite modélisés sous forme de Complexes Segmentaires Inter Points. Différentes modélisations 3D ont été réalisées: modélisation de l’os pariétal avec 9 points de repères craniométriques (figure 2), et de la voûte bipariétale au moyen de 15 points de repères. En présence d’os pariétaux isolés, nous avons symétrisé l’os par rapport au plan défini par la suture sagittale (figure 3, exemple de la reconstruction virtuelle du crâne Arago 21-47). Dans chaque cas, les modélisations des différents fossiles ont subi une superimposition Procuste et ont été analysées au moyen du logiciel Morphologika ®.
Résultats
Comparaison Procuste du pariétal droit (figure 4)
L’axe 1 de l’ACP permet d’expliquer 20,5% de la variance totale de l’effectif fossile et moderne étudié, contre 14,2% pour l’axe 2.
Le passage des valeurs négatives de l’axe 1 (Sts 5; Australopithecus africanus) aux valeurs positives (Homo sapiens historique) correspond à une réduction de la longueur antéro-postérieure de l’os pariétal et à un accroissement de la hauteur de l’écaille. Ceci correspond à un changement de courbure de l’écaille pariétale, avec un sagittum plus élevé par rapport à l’axe bregma-lambda (avec un lambda plus bas). L’astérion est positionné plus antérieurement par rapport au lambda et plus bas par rapport au sphénion et au krotaphion, tout comme l’incisure pariétale.
Le passage des valeurs négatives de l’axe 2 (Homo sapiens historique) aux valeurs positives (Homo neanderthalensis) correspond à une position plus postérieure de la bosse pariétale (ou zone assimilée) par rapport au bord coronal, plus haute et plus interne par rapport à la modélisation de la suture pariéto-temporale.
Nous ne pouvons distinguer les phylums les uns des autres à l’exception des Néandertaliens, correspondant à l’extrémité positive de l’axe 2.
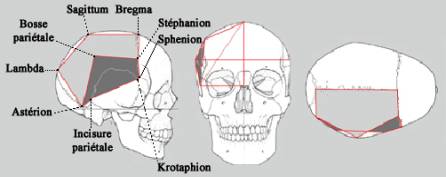
Figure 2: Modélisation de l’os pariétal droit au moyen de points de repères craniométriques.

Figure 3: Obtention virtuelle par symétrie sagittale du pariétal gauche à partir du pariétal droit Arago 47.
Comparaison Procuste de la voûte bipariétale (figure 4)
L’axe 1 de l’ACP permet d’expliquer 23,9% de la variance totale de la population contre 11,5% pour l’axe 2. Ceci correspond à une plus grande valeur discriminative de l’axe 1.
Le passage des valeurs négatives de l’axe 1 (Sts 5; Australopithecus africanus) aux valeurs positives (Homo sapiens historique) correspond à une réduction de la longueur antéro-postérieure de la voûte bipariétale et à un accroissement de sa hauteur. Ceci correspond à un changement de courbure de la voûte bipariétale, avec un sagittum plus élevé par rapport à l’axe bregma-lambda (avec un lambda plus bas). Les astérions sont positionnés plus antérieurement par rapport au lambda et plus bas par rapport aux sphénions et aux krotaphions (tout comme les incisures pariétales). Les bosses pariétales sont aussi positionnées de manière plus externe. Ceci correspond à un bombement de la voûte bipariétale dans le plan frontal.
Le passage des valeurs négatives de l’axe 2 (Sts 5; Australopithecus africanus) aux valeurs positives (Homo neanderthalensis) correspond à une position plus postérieure de la bosse pariétale (ou zone assimilée) par rapport au bord coronal, plus haute et plus interne par rapport à la modélisation de la suture pariéto-temporale. Ceci traduit une différence de répartition des largeurs maximales de la voûte bipariétale.
Les phylums peuvent être mieux discriminés les uns par rapport aux autres. Nous ne pouvons cependant pas ou peu distinguer les Homo sapiens préhistoriques des individus historiques.
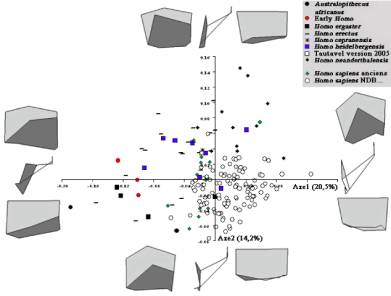
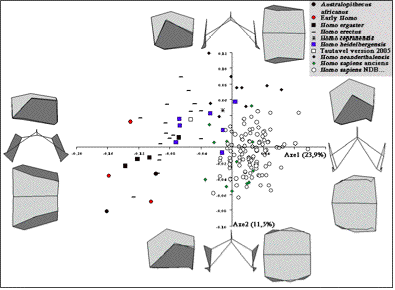
Figure 4: Analyses en composantes principales des modélisations de l’os pariétal droit et de la voûte bipariétale, en projection dans le plan latéral, frontal et supérieur.
Discussion
L’étude de la configuration tridimensionnelle de l’os pariétal isolé permet de distinguer morphologiquement certaines populations fossiles telles que les Néandertaliens, mais la discrimination des différents phylums à partir de ces seules données pariétales est médiocre. Il est ainsi impossible de distinguer les Homo heidelbergensis des Homo sapiens préhistoriques. Par contre, la comparaison des voûtes bipariétales permet de mieux distinguer ces différents phylums. Nous pouvons ainsi définir plus clairement des conformations consensus pour certaines espèces (Homo ergaster, Homo heidelbergensis, Homo erectus, Néandertaliens et Homo sapiens).
La discrimination accrue entre les phylums dans le cadre de la modélisation de la voûte bipariétale par rapport à la modélisation de l’os pariétal droit isolé est due à la présence de nouvelles angulations. De nouveaux angles apparaissent entre les points de repères latéraux et doivent être pris en compte. Ceux-ci permettent d’accentuer la précision de la modélisation et des données obtenues par ACP. Ainsi, alors que l’un des deux Australopithecus africanus modélisé présente une conformation spatiale de l’os pariétal droit comparable à celle d’Homo sapiens historiques, la discrimination entre ces individus est nette lors de l’analyse de la voûte bipariétale.
Cet exemple illustre la nécessité de multiplier les modélisations tridimensionnelles différentes, même si l’analyse d’une seule modélisation apporte déjà des informations conformationnelles et permet de discriminer certains phylums.
Conclusion
Le pariétal est un os de la voûte du crâne souvent bien conservé. Son étude 3D de manière isolée par méthode Procuste ne permet pas une discrimination nette des phylums des Hominidés, montrant ainsi les limites de son étude en tant qu’os isolé. La reconstruction virtuelle de la voûte bipariétale est possible par symétrisation sagittale du côté conservé et peut éventuellement être associée à une compensation des déformations du frontal et/ou de l’occipital pour préciser l’angle lambdatique. Les analyses Procuste réalisées à partir de ces voûtes sont nettement plus discriminantes.
L’utilisation de l’imagerie 3D pour la reconstitution des os manquants par symétrisation (position spatiale relative; Guipert et al., 2003, 2004; Guipert, 2005), permet d’obtenir d’avantages de données ostéologiques et donc d’optimiser les analyses morphométriques.
Bibliographie
Bookstein, F. L., 1989, Principal warps: thin-plate splines and the decomposition of deformations. IEEE Trans. Pattern Analysis Machine Intelligence, 11, 567-585.
Bookstein, F. L., 1991, Morphometric tools for landmark data. Geometry and biology (Cambridge: Cambridge University Press).
Bookstein, F. L., 1996, Combining the tools of geometric morphometrics. Dans Advances in Morphometrics édité par L.F.M. et al. (New York: Plenum Press), pp. 131-151.
Bookstein, F. L., 1997, Landmark methods for forms without landmarks: morphometrics of group differences in outline shape. Medical Image Analysis, 1, 225-243.
Bookstein, F., Schäfer, K., Prossinger, H., Seidler, H., Fieder, M., Stringer, C., Weber, G. W., Arsuaga, J-L., Slice, D. E., Rohlf, F. J., Recheis, W., Mariam, A. J., et Marcus, L. F., 1999, Comparing Frontal Cranial Profiles in Archaic and Modern Homo by Morphometric Analysis. The Anatomical Record (New Anat.), 257, 217-224.
Bookstein, F. L., Gunz, P., Mitteroecker, P., Prossinger, H., Schaefer, K., et Seidler, H., 2003, Cranial integration in Homo: singular warps analysis of the midsagittal plane in ontogeny and evolution. Journal of Human Evolution, 44, 167-187.
Delattre, A., Fénart, R., 1960, L’hominisation du crâne étudiée par la méthode vestibulaire. Edité par CNRS Editions (Paris).
Delson, E., Harvati, K., Reddy, D., Marcus, L. F., Mowbray, K., Sawyer, G. J., Jacob, T., et Marquez, S., 2001, The Sambungmachan 3 Homo erectus Calvaria: A Comparative Morphometric and Morphological Analysis. The Anatomical Record (New Anat.), 262, 380-397.
Grimaud-Hervé, D, Jacob, T., 1983, Les pariétaux du Pithécanthrope Sangiran 10. L’Anthropologie (Paris), 87, 469-474.
Guipert, G., 2005, Reconstitution et position phylétique des restes crâniens de l’Homme de Tautavel (Arago 21-47) et de Biache-Saint-Vaast 2. Apports de l’imagerie et de l’analyse tridimensionnelles. Thèse de Doctorat, Université Paul Cézanne Aix-Marseille III, pp. 480.
Guipert, G., Subsol, G., Jessel, J.-P., Delingette, H., et Mafart, B., 2003, The Fovea Project: A New Look at Human Past. Dans Proceedings of the VSMM 2003, édité par VSMM, pp. 43-47.
Guipert, G., Lumley, M.-A. de, Lumley, H. de, et Mafart, B., 2004, Three-Dimensional Imagery: a New Look at the Tautavel Man. Dans [ Enter the Past ] The E-way into the Four Dimensions of Cultural Heritage | CAA 2003 | Computer Applications and Quantitative Methods | Proceedings of the 31st Conference, Vienna, Austria, April 2003, édité par SW (Wien: BAR International Series) pp. 100-102.
Guipert, G., et Mafart, B., sous presse, Evolution de l’os frontal d’Homo heidelbergensis et Homo neanderthalensis et comparaison avec l’Homme moderne. Anthropologie (Brno).
Harvati, K., 2002, Models of Shape Variation between and within species and the Neanderthal Taxonomic position: A 3D Geometric Morphometrics Approach Based on Temporal Bone Morphology. Dans Three-Dimensional Imaging in Paleoanthropology and Prehistoric Archaeology, édité par Mafart, B., et Delingette, H. (Oxford: Archaeopress, Publishers of British Archaeological Reports) pp. 25-30.
Harvati, K., 2003, Quantitative Analysis of Neanderthal Temporal Bone Morphology Using Three-Dimensional Geometric Morphometrics. American Journal of Physical Anthropology, 120, 323-338.
Mafart, B., 2002, 3D Imaging in Paleoanthropology and Prehistoric Archeology: A New Tool for Old Sciences or an Emerging Science? Dans Three-Dimensional Imaging in Paleoanthropology and Prehistoric Archaeology, édité par Mafart, B., Delingette, H. (Oxford: Archaeopress, Publishers of British Archaeological Reports) pp. 1-3.
Mafart, B., Guipert, G., Lumley, M.-A. de, Subsol, G., 2004, Three-dimensional computer imaging of hominid fossils: a new step in human evolution studies. Canadian Association of Radiologist Journal, 55, 264-270.
Martin, R., Saller, K., 1957, Lehrbuch der Anthropologie in Systematischer Darstellung mit Besonderer Berücksichtigung der Anthopologischen Methoden (Stuttgart: Gustav Fischer Verlag), pp. 661.
Penin, X, et Baylac, M., 1995, Analysis of the skull shape changes in Apes using 3D Procustes superimposition. Dans Current Issues in Statistical Shape Analysis, édité par Mardia, K., V., Gill, C., A. (Leeds: Leeds University Press) pp. 208-210.
Zollikofer, C. P. E., 2002, A Computational Approach to Paleoanthropology. Evolutionary Anthropology, 11, 64-67.
Zollikofer, C. P. E., et Ponce de Leon, M. S., 2002, Virtual Paleoanthropology: the 4th Dimension. Dans Three-Dimensional Imaging in Paleoanthropology and Prehistoric Archaeology, édité par Mafart, B., Delingette, H. (Oxford: Archaeopress, Publishers of British Archaeological Reports) pp. 47-53.