
Colloques
du Groupement des Anthropologistes de Langue Française (GALF)
Jeusel, E., Mafart, B., 2006, Le palais des hominidés. Comparaison avec les grands singes et l’homme moderne. Résultats préliminaires. Antropo, 11, 235-247. www.didac.ehu.es/antropo
Le
palais des hominidés. Comparaison avec les grands singes et l’homme moderne.
Résultats préliminaires
Hominids’s
hard palate. Comparison between apes and modern humans. Preliminary results
Elise Jeusel, Bertrand Mafart
Antenne
de l’Institut de Paléontologie Humaine, CNRS-CEREGE Université Paul Cézanne
(Aix-Marseille III), Europôle Méditerranéen de l’Arbois, BP 80 13 345
Aix-en-Provence. E-mail: elise.jeusel@caramail.com
Mots-clés: cavité palatine, empreintes,
3D, hominidés, grands singes.
Key-words: palatine cavity, casts, 3D,
hominids, apes.
Résumé
Notre étude porte sur l’évolution morphométrique et la modélisation de la cavité palatine des hominidés dans la perspective de l’acquisition d’un langage articulé. Nous avons étudié le palais de 50 hommes modernes, 6 chimpanzés, 9 gorilles, et 7 hominidés fossiles. Des moulages en silicone de la cavité palatine avec comme limite postérieure le plan frontal passant par les deuxièmes molaires supérieures ont été analysés au plan morphologique et anatomique. Des superpositions Procrustes et des analyses en composantes principales (logiciel Morphologika©) ont été réalisées à partir des coordonnées tridimensionnelles des points caractéristiques de la cavité palatine, relevés avec un digitaliseur Microscribe® 3DX. Nous avons pu caractériser la configuration de l’homme moderne dont le palais est court, large, et relativement profond, et la configuration des grands singes dont le palais est long, étroit, et peu profond. Le palais d’un australopithèque AL 200-1 (Australopithecus afarensis) a une configuration proche de celle des chimpanzés. Cependant, ce fossile présente un rapport largeur/hauteur allant des P3/ aux M2/ qui le distingue à la fois des grands singes et des hominidés fossiles et modernes. Le palais osseux des espèces fossiles du genre Homo présente une variation morphologique relativement faible avec celui de l’homme moderne. Nos résultats montrent que les australopithèques n’avaient donc probablement pas un palais permettant la production d’un langage articulé. Une morphologie du palais osseux compatible avec cette fonction semble présente dès les plus anciens représentants du genre Homo. Cependant, les éléments non osseux du complexe anatomique n’étant pas conservés, il n’est pas possible de conclure plus avant actuellement.
Abstract
Our study relates to the morphometrical evolution and the modelisation of the palatine cavity of hominids in the prospect of the acquisition of an articulated language. We studied the hard palate of 50 modern humans, 6 chimpanzees, 9 gorillas, and 7 fossil hominids. Silicon casts of the palatine cavity, with as posterior limit the frontal plan passing by the second upper molars, were analyzed morphologically and anatomically. Procrustes superpositions and principal component analysis (Morphologika© Freeware) have been realized from the three-dimensional co-ordinates of the point characteristics of the palatine cavity, collected with a digitizer Microscribe® 3DX. We could characterize the configuration of the modern human whose palate is short, broad, and relatively deep, and the configuration of the apes whose palate is long, narrow, and not very deep. The AL 200-1 palate (Australopithecus afarensis) has a configuration close to that of the chimpanzees. However, this fossil presents a width/height ratio going from P3/ to M2/ which distinguish it from both apes and fossil and modern hominids. The hard palate of fossil species of the genus Homo presents a relatively weak morphological variation with the modern human’s one. Our results show that the Australopithecus did probably not have a palate allowing the production of an articulated language. Morphology of the hard palate compatible with this function seems present from the earliest fossils of the genus Homo. However, the soft elements of the anatomical complex not being preserved, we can’t conclude any farther at this time.
Introduction
Le langage articulé est une des
spécificités de l’Homme au sein du règne animal. Il
résulte de capacités cognitives et d’une conformation anatomique particulière
dont les mécanismes sont considérés comme
connus. Depuis longtemps, les paléoanthropologues s’intéressent à la question
du moment de l’apparition du langage articulé au cours de l’évolution humaine,
en étudiant d’une part l’aspect cognitif, et d’autre part la morphologie et la
biomécanique du tractus respiratoire supérieur (ou tractus vocal). Ainsi, à
partir de la position relative du tractus par rapport aux structures osseuses
qui le soutiennent chez les hommes modernes et les grands singes, plusieurs
reconstitutions du tractus vocal des hominidés fossiles ont été proposées,
notamment pour les australopithèques (Laitman, 1986 et 1985; Laitman et
Heimbuch, 1982; Lieberman 1975 ), pour les néandertaliens (Arensburg et
Tillier, 1991; Laitman et al., 1991; Arensburg et al., 1990 et 1989; Lieberman,
1975), et plus récemment sur la
base d’une nouvelle analyse anatomique (Heim et al., 2000 et 2002; Boë et
al., 2002).
Malheureusement, la majeure partie du tractus vocal, essentiellement constituée
de tissus non osseux, n’est pas fossilisée. Seul
le palais osseux est conservé mais rarement complet en raison de la fragilité
des os qui le constituent.
Classiquement, l’aspect cognitif de la parole est considéré comme reposant sur la notion d’asymétrie du cortex cérébral (Deacon, 1992; Bradshaw et Rogers, 1993). Chez l’homme, il existe deux zones principales correspondant aux aires associatives du langage situées dans l’hémisphère gauche: l’aire de Broca, impliquée dans la composante motrice du langage (production et articulation), et l’aire de Wernicke, intervenant dans la perception des mots et leur compréhension. Des travaux récents de neuroanatomie et neurophysiologie ont montré que les relations neuroniques lors de la production du langage sont extrêmement complexes et ne sont pas limitées à ces deux zones corticales (Habib et al., 2000; Pulvermuller, 2002, Lieberman, 2002). Ainsi, l’ensemble de la zone entourant la scissure de Sylvius serait spécialisé dans différents aspects du langage, et la notion d’asymétrie serait à relativiser puisque les seules aires associatives ne sont pas les seules à être latéralisées dans le cerveau humain (Habib et al., 2000). Les recherches sur les primates non-humains se heurtent à la difficulté de réalisation d’études neurophysiologiques aussi complexes que pour l’homme. Des travaux récents auraient localisé une aire similaire à l’aire de Broca chez les genres Pan et Gorilla (Sherwood et al., 2003; Cantalupo et Hopkins, 2001) et une aire similaire à l’aire de Wernicke chez les grands singes (Gannon et al., 1998; Hopkins et al., 1998). Les connaissances des possibilités cognitives de ces primates restent cependant très limitées.
L’architecture de base du tractus respiratoire supérieur, commune à tous les mammifères, comprend le larynx, la cavité buccale constituée par le palais et la mandibule, et l’orifice buccal. De nombreux travaux ont été consacrés à l’étude de la position du larynx (Nishimura et al., 2003; Boë et al., 2002; Fitch et Reby, 2001; Heim et al., 2000; Falk, 1975). Le larynx du nouveau-né a une position haute qui évolue vers l’âge de deux ans, vers une position basse (en regard de la quatrième vertèbre cervicale chez l’homme (Heim et al., 2002)). Les primates non-humains conservent au cours de leur vie adulte un larynx haut (Lieberman, 1992) (Figure 1).
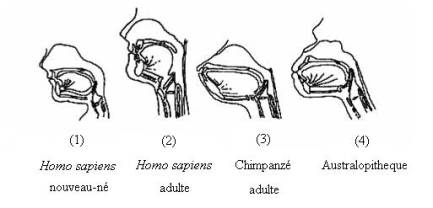
Figure 1.
Schématisation des tractus vocaux d’un nouveau-né humain, d’un homme adulte,
d’un chimpanzé adulte, et d’un australopithèque. (1): modifié d’après Laitman,
Crelin, et Conlogue, 1977; in Lieberman, 1984; (2): modifié d’après Lieberman,
1984; (3) et (4): modifiés d’après Laitman et Heimbuch, 1982; in Lieberman,
1984.
Figure 1. Vocal
tracts schematization of a human newborn, an adult man, an adult chimpanzee,
and an australopithecine. (1): modified after Laitman, Crelin, et Conlogue,
1977; in Lieberman, 1984; (2) modified after Lieberman, 1984; (3) et (4):
modified after Laitman et Heimbuch, 1982; in Lieberman, 1984.
Différents auteurs réfutent l’idée de langage articulé lié à la descente du larynx des adultes humains en faisant appel à l’anatomie d’autres espèces animales (Fitch et Reby, 2001). La descente du larynx, s’effectuant à la puberté chez l’homme adulte, n’augmenterait pas les capacités phonétiques mais augmenterait la longueur du tractus vocal, baissant ainsi les fréquences formantes. La descente du larynx ne serait pas directement liée à la parole et ne constituerait qu’une préadaptation.
D’un autre côté, Nishimura et al. (2003) proposent une évolution du larynx dans la lignée des hominoïdes et non pas seulement dans la lignée des hominidés sur la base de l’observation de jeunes chimpanzés. Au cours de leur développement, ils ont observé une descente du larynx similaire aux jeunes enfants, qu’ils interprètent comme une préadaptation aux modifications fonctionnelles conduisant à l’évolution du langage articulé des hommes modernes. L’intérêt de l’étude de la morphologie du larynx dans l’analyse de la parole est ainsi remis en question.
L’aptitude à produire un langage articulé dépend également de la forme du tractus vocal (Tattersall, 1998). Les grands singes possèdent un tractus vocal apte à la production de certains sons différenciés (Zimmermann, 1992), permettant un certain niveau de communication entre eux mais inadapté au langage articulé. De nombreuses études sur l’apprentissage du langage chez les chimpanzés ont montré qu’ils sont capables de comprendre des phrases assez simples mais qu’en aucun cas ils ne possèdent un langage articulé (Savage-Rumbaugh, 1992).
La possibilité d’un langage articulé dépend aussi de la
capacité de la langue d’atteindre certaines zones du tractus vocal supérieur et
notamment du palais. Certains auteurs (Kay et al., 1998) avaient émis l’hypothèse d’une
corrélation entre la taille du nerf hypoglosse, innervant entre autre la
langue, et la complexité de la musculature de la celle-ci, et prêtaient ainsi aux australopithèques des
capacités langagières nettement supérieures aux grands singes non justifiées
par la découverte d’artéfacts culturels associés. Cette hypothèse a été
récemment rejetée par DeGusta et al. (1999) et Jungers et al. (2003) qui ont démontré que
la taille du nerf hypoglosse ne permet pas de distinguer les chimpanzés des
humains modernes.
Les données anatomiques et cognitives sur les hominidés actuels nous renseignent sur les pré-requis à un langage articulé. Les données fossiles, naturellement restreintes, ne nous donnent que de faibles indices concernant leurs capacités.
L’étude du palais osseux, dans la perspective de la production d’un langage articulé, est donc essentielle compte tenu de la disparition des parties molles. Contrairement à l’os hyoïde, le palais osseux est fixe et fait partie intégrante du crâne. Malheureusement, les palais d’hominidés fossiles et même de séries de comparaisons historiques sont souvent incomplets et/ou déformés. Les principaux problèmes rencontrés sont une mauvaise conservation post-mortem (absence de certaines dents, fractures, déformations), et un état « biologique » limitant l’analyse (pathologie, morphologie liée à l’âge), ainsi que la relative asymétrie naturelle du palais.
Le palais est une structure anatomique formant une cloison entre les cavités nasales et buccale. Il est composé d’une partie osseuse et d’une partie constituée de tissus mous. La partie osseuse ou palais dur est composée par les deux processus palatins du maxillaire et les deux apophyses horizontales des os palatins (Paturet, 1951), ainsi que par le prémaxillaire chez les jeunes enfants et les grands singes même adultes (Swindler et Wood, 1973). La jonction de ces os forme la suture médio-palatine et la suture palatine transverse (Le Diascorn, 1972). La partie molle, ou voile du palais, est implantée à l’arrière du palais dur et se termine par la luette. Ces deux parties sont recouvertes d’une fine muqueuse. La voûte ainsi constituée est plus ou moins concave vers le bas dans le plan sagittal et dans le sens transversal selon les espèces de primates considérées. Le palais de l’homme moderne est court, large, et profond dans la grande majorité des cas et présente de petits reliefs et dépressions. A l’intérieur de la cavité buccale, la langue, extrêmement mobile chez l’homme, utilise au mieux l’espace disponible. Le palais des grands singes est long, étroit, et peu profond avec une surface lisse, et la langue comble la totalité de l’espace buccal, tout comme chez le nouveau-né humain (Lieberman, 1975).
La production du langage articulé ne
dépend pas uniquement, loin s’en faut, de la seule morphologie du palais osseux
(Tattersall, 1998; Habib et al., 2000; Heim et al., 2000; Boë et al., 2002; Heim et al., 2002; Lieberman,
2002; Pulvermuller, 2002; Zhang et al., 2002; Clark et al., 2003; Marcus et al., 2003; Takahashi et al., 2003). Cependant, la
production de sons dépend de facteurs indirectement liés à la profondeur du
palais. Ainsi, l’existence d’une langue capable de mouvements complexes (Kay et
al., 1998;
DeGusta, 1999; Jungers et al., 2003), organe essentiel pour la production de sons
complexes, est liée à la morphologie palatine.
Matériel
50 crânes secs d’Homo sapiens modernes, 6 de chimpanzés (Pan
sp.) et 9 de gorilles (Gorilla sp.) ont été examinés. Les moulages en
résine des palais osseux de 7 hominidés fossiles suffisamment bien conservés
ont été étudiés (Tableau 1). Les palais devaient
pouvoir être décris d’un point de vue anatomique et comporter les points de
repère choisi pour l’étude tridimensionnelle.
Des empreintes de la cavité palatine ont
été faites avec du polysiloxane (silicone par addition). Ces empreintes ont été
digitalisées avec le MicroScribe® 3DX. Les coordonnées 3D ont été traitées avec
le Logiciel Morphologika© (Freeware).
|
N° inventaire |
Attribution
systématique |
Localité |
Age absolu (BP) |
Moulage |
Références |
|
AL 200-1 |
Australopithecus
afarensis |
Hadar, Ethiopie |
3,0 Ma +/- 0,2 |
M.Taieb |
Johanson et Taieb,
1976, Taieb et al., 1976, Johanson et White,
1979 |
|
OH 5 |
Paranthropus boisei |
Olduvai, Tanzanie |
1,75 Ma |
F. Wenner-Gren |
Leakey, 1959, Tobias, 1967 |
|
KNM- WT 15 000 A |
Homo ergaster |
Nariokotome, Kenya |
1,6 Ma |
R. Leakey |
Brown et al., 1985, Walker et Leakey, 1993 |
|
Broken Hill 1 (Kabwe) |
Homo sp. Attribution incertaine |
Kabwe, Zambie |
-300 000/ -200 000 ans |
F. Wenner-Gren |
Vrba, 1982, Partridge, 1982 |
|
Qafzeh 6 |
Homo sapiens |
Qafzeh, Israël |
- 100 000 / - 90 000 ans |
Y. Rak |
Vandermeersch,
1981, Valladas
et al., 1991, Yokoyama
et al., 1997 |
|
Qafzeh
9 |
Homo
sapiens |
Qafzeh,
Israël |
-
100 000 / -
90 000 ans |
Y.
Rak |
Vandermeersch,
1981, Valladas et al., 1991, Yokoyama et al., 1997 |
|
La Ferrassie 1 |
Homo
neanderthalensis |
Abri de La Ferrassie,
France |
0,072 Ma |
Musée de l’Homme,
Paris |
Heim, 1976, Mellars, 1986 |
Tableau 1. Hominidés
fossiles étudiés.
Table 1. Fossil
hominids studied.
Méthode
Les individus sélectionnés
sont adultes et exempts de pathologie. Les arcades dentaires comportent au
moins les dents de repère, le palais est en bon état, et non déformé.
Des empreintes en silicone ont été prises de la cavité palatine avec pour limite postérieure, les collets internes des M2/ (Figure 2). Un plan de référence a été choisi, passant par le point inter-incisif médian, et les collets des M2/ droite et gauche. Les empreintes sont préparées selon un protocole détaillé dans une étude précédente (Jeusel et Mafart, 2003). Ces empreintes font l’objet d’une étude de volume, d’une étude métrique, d’une étude anatomique (Figure 3), ainsi que d’une étude tridimensionnelle.

Figure 2. Empreinte
en silicone d’un palais d’homme moderne.
Figure 2. Silicon
cast of a modern human palate.
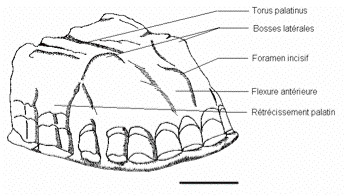
Figure 3.
Schématisation d’une empreinte en silicone et quelques structures anatomiques
modernes.
Figure 3.
Schematization of a silicon cast and some modern anatomical structures.
A partir des empreintes, nous avons obtenu 4 profils
frontaux, 1 profil sagittal, et 1 triangle basal (plan de référence) (Figure
4). A partir de ces profils, nous avons obtenu une série de dimensions de la cavité palatine.
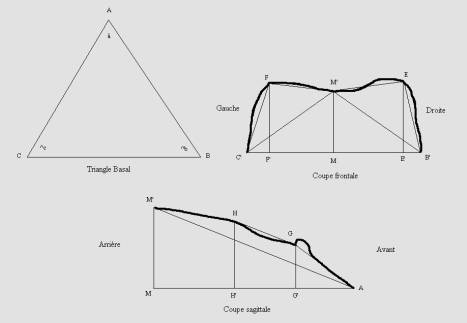
Figure 4. Unités
géométriques définies sur les empreintes en silicone.
Figure 4. Geometrical
units determinated from the silicone casts.
A partir de ces mêmes empreintes, les points de repère ont été digitalisés avec le MicroScribe® 3DX. Les coordonnées ainsi obtenues, ont été analysées avec le logiciel freeware Morphologika© (Figure 5).
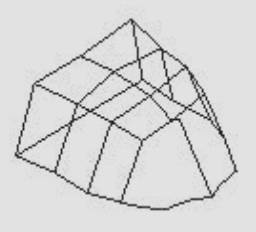
Figure 5.
Modélisation tridimensionnelle avec le logiciel Morphologika© d’un palais
d’homme moderne.
Figure 5.
Morphologika© freeware three-dimensional modelisation of a modern hard palate.
Résultats préliminaires
Les grands singes ont une
largeur palatine quasi-constante des M2/ aux P3/, alors que celle des hommes
modernes diminue fortement (Figure 6).
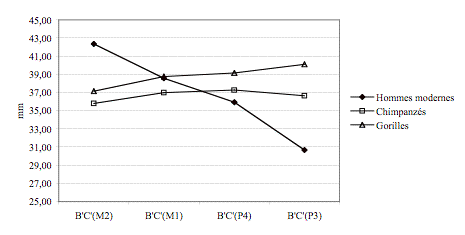
Figure 6. Evolution
de la largeur palatine en fonction de la dent considérée.
Figure 6. Palatine
width evolution according to the considered tooth.
Les hauteurs palatines diminuent des M2/ aux P3/ quelque
soit l’espèce considérée (Figure 7). Les hauteurs des hommes modernes se
situent entre celles des gorilles et celles des chimpanzés.
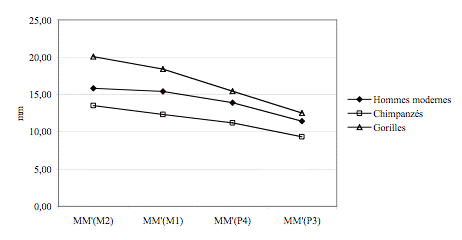
Figure 7. Evolution
de la hauteur palatine en fonction de la dent considérée.
Figure 7. Palatine
height evolution according to the considered tooth.
La comparaison des trois dimensions principales de la cavité palatine des différents hominidés et grands singes, montre que AL 200-1 à des dimensions similaires aux chimpanzés (Figure 8). Les dimensions de KNM-WT 15 000 A sont intermédiaires entre chimpanzés et hommes modernes. Tous les autres hominidés fossiles récents ont des dimensions plus proches de celles des hommes modernes.
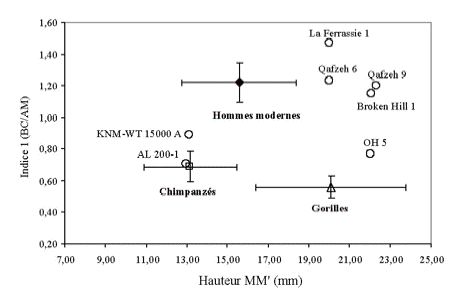
Figure 8. Comparaison
des trois dimensions principales palatines.
Figure 8. Comparison of the three principal palatine dimensions.
L’indice 3 (BC/MM’) nous montre que le palais de AL 200-1 se singularise par rapport aux autres hominidés et grands singes (Figure 9). La profondeur palatine antérieure est très faible alors que sa largeur est quasi-constante des M1/ aux P3/.
|
|
|
|
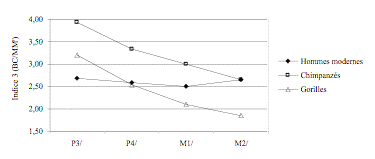
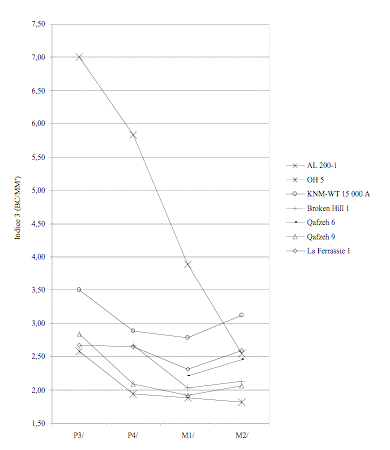
Figure 9. Evolution
de l’indice 3 en fonction de la dent considérée. (a) Hommes modernes et grands
singes. (b) Hominidés fossiles.
Figure 9. Evolution
of the indice 3 according to the considered tooth. (a) Modern humans and apes.
(b) Fossil hominids.
Discussion
La méthode utilisant des moulages en silicone
permet de visualiser le palais en tant que cavité fonctionnelle. Ces moulages
permettent également d’avoir accès à un grand nombre de dimensions. Cependant,
l’emploi de la silicone ne peut se faire directement sur un fossile original du
fait de sa fragilité. Cette étude doit donc être complétée par des observations
anatomiques sur les fossiles originaux, permettant de vérifier la validité de
la reconstruction utilisée.
L’étude morphométrique des empreintes en silicone du palais osseux a permis néanmoins d’évaluer les variations inter-spécifiques et intra-spécifiques des hommes modernes, des chimpanzés, des gorilles, et de quelques hominidés fossiles (Jeusel et Mafart, 2003).
La conformation de type moderne est observée, avec des variations minimes, chez tous les représentants récents du genre Homo étudiés.
Il
est apparu que globalement, les hominidés fossiles anciens (australopithèques
et paranthropes) ont une morphométrie très proche de celle des grands singes
avec une cavité palatine longue et étroite. Le palais de AL 200-1 (Australopithecus
afarensis) a
une configuration globalement simienne. Cependant, le palais osseux du fossile
est très peu profond dans sa partie antérieure et se distingue à la fois des
grands singes et du genre Homo. Bien que la production du langage articulé implique
d’autres structures anatomiques, rarement fossilisées, la morphologie palatine
est en défaveur de l’existence d’un langage articulé pour cet hominidé. Il est
donc vraisemblable que l’absence de profondeur du palais de l’australopithèque
est un élément anatomique défavorable à la production d’un langage articulé
(Arnold et al.,
2004).
Laitman et Heimbuch (1982) et Laitman (1985), par leurs études menées sur le tractus respiratoire supérieur des australopithèques, a considéré l’association d’une flexion basicrânienne très faible ainsi qu’une cavité buccale longue, remplie par une langue volumineuse et peu mobile, comme une morphologie proche des grands singes. Le larynx se trouverait ainsi en position haute. Pour cet auteur, les australopithèques ne possédaient pas de langage articulé. Ils n’auraient pas été capables de produire la gamme de sons caractéristique du langage humain (Leakey, 1994). Lieberman (1975) attribuait au fossile Sterkfontein 5 (Australopithecus africanus), les mêmes limitations phonétiques que le chimpanzé au vu de la reconstruction de son tractus vocal. Sur le plan cognitif, les aires du langage ne seraient pas bien individualisées (De Lumley, 1998).
L’Homo habilis/rudolfensis est représenté par très peu de
crânes complets. Les deux spécimens les plus complets sont KNM-ER 1813 et
KNM-ER 1470. Tous deux sont déformés. Le palais du premier ressemble aux
australopithèques, le palais du second est extrêmement altéré. L’étude du
cortex cérébral de ces hominidés ferait apparaître des prédispositions au
langage (Deacon, 1992), corroborées par l’apparition d’outils manufacturés qui
semble indiquer la nécessité d’une forme de communication élaborée, peut-être
pas articulé comme l’homme moderne (Reeves et al., 1996).
L’Homo
ergaster/erectus a
été peu étudié puisque la majorité des restes fossiles retrouvés ne comporte
pas de palais. La nécessité d’une forme de communication élaborée semble
indiscutable compte tenu de son évolution technologique, de son habitat et de
ses habitudes de chasse (De Lumley, 1998) mais encore difficile à prouver.
Laitman (1984; 1985), sur la base de ses travaux sur la flexion du basicrâne de
fossiles africains, situe la restructuration du tractus respiratoire supérieur
chez ces hominidés. Une flexion minime semble indiquer le début de la descente
du larynx, changeant ainsi le modèle respiratoire basique des mammifères en
modèle humain. Les études menées sur les moulages endocrâniens de quelques
spécimens d’Homo erectus/ergaster ont démontré la présence d’une certaine
asymétrie du cortex cérébral (Broadfield et al., 2001; Holloway, 1982). Cependant, le
fossile KNM-WT 15 000 A ne semble pas avoir un palais permettant l’articulation
complète.
Bénéficiant d’une structure
sociale développée, de techniques de fabrication d’outillage élaborées, d’une
capacité crânienne importante, il est difficilement concevable que Homo
neanderthalensis
ne dispose pas d’un langage complexe. Pourtant, sur la base de la première
reconstitution du crâne de La Chapelle-aux-Saints faite par Boule et Papoint
(1911-1913), Lieberman (1975) ne lui attribuait pas la capacité de parler,
positionnant le larynx haut dans le cou. Laitman et al. (1991; 1996), quant à eux,
expliquent la différence de morphologie de la zone laryngée par l’adaptation spécifique à un environnement
différent, précisant que le tractus respiratoire supérieur joue aussi un rôle
dans les fonctions respiratoires et digestives. La flexion basicrânienne,
moindre que les Homo ergaster/erectus, n’aurait pas forcément beaucoup affecté
la capacité langagière de cette espèce (Leakey, 1994). Récemment, l’approche
phonétique aurait permis de conclure que même avec un larynx en position haute,
la conformation de leur tractus
vocal leur aurait permis de produire des voyelles aussi différenciées que
celles de l’homme moderne (Boë et al., 2002; Heim et al., 2002).
Notre étude confirme que les
hommes de Neandertal avaient probablement un tractus supérieur apte à la
prononciation du langage articulé.
La morphologie palatine
moderne semble être acquises chez les Homo sapiens archaïques qui possèdent une
anatomie moderne du tractus vocal (Lieberman, 1975) et du cortex cérébral. Rien
ne les distingue véritablement des hommes actuels. Leur évolution culturelle et
technologique semble être la preuve de l’existence d’un langage articulé vrai.
Laitman (1984 et 1985) situe la flexion complète du basicrâne chez ces
hominidés.
Le palais de Broken Hill 1 est
d’apparence très moderne. Son seul palais pourrait le faire entrer dans
l’espèce moderne. Ses caractères crâniens archaïques semblent indiquer une
évolution différenciée du palais.
Un palais moderne est donc en
place chez les hominidés récents tels que Broken Hill 1, Qafzeh 6 et 9, et La
Ferrassie 1, qui réunissent toutes les structures anatomiques permettant la
production d’un langage articulé de type moderne.
L’étude du palais osseux des hominidés fossiles apporte des arguments pour déterminer l’apparition de la première morphologie palatine ayant pu permettre la production d’un langage articulé. Cependant un palais anatomiquement moderne n’implique pas la production d’un langage articulé mais seulement l’aptitude morphologique à produire ce langage.
Cette étude se poursuit
actuellement en incluant d’autres hominidés fossiles.
Bibliographie
Arensburg, B., Tillier, A.M.,
1991, Speech and the Neanderthals. Endeavour, 15, 26-28.
Arensburg, B., Schepartz,
L.A., Tillier, A.M., Vandermeersch, B., Rak, Y., 1990,
A reappraisal of the anatomical basis for speech in Middle Palaeolithic
hominids, American Journal of Physical Anthropology, 83, 137-146.
Arensburg, B., Tillier, A.M.,
Vandermeersch, B., Duday, H., Schepartz, L.A., Rak, Y., 1989, A Middle
Palaeolithic human hyoid bone, Nature, 338, 758-760.
Arnorld, W.H., Zoellner, A.,
Sebastian, T., 2004, Development of the palatal size in Pan troglodytes, Hominids and Homo sapiens. Annals of Anatomy, 186,
511-520.
Boë, L.-J., Heim, J.-L.,
Honda, K., Maeda, S., 2002, The potential Neandertal vowel space was as large
as that of modern humans. Journal of Phonetics, 30, 465-484.
Bradshaw, J. L., Rogers L. J.,
1993, The evolution of lateral asymmetries, language, tool use, and intellect.
(San Diego: Harcourt Brace Jovanovich, Publishers, Academic Presse Inc.), pp.
463.
Broadfield,
D.C., Holloway, R.L., Mowbray, K., Silvers, A., Yuan, M.S., Marquez, S., 2001,
Endocast of Sambungmacan 3 (Sm 3): A New Homo erectus From Indonesia. The
Anatomical Record, 262, 369-379.
Brown,
F., Harris, J., Leakey, R., Walker, A., 1985, Early Homo erectus skeleton from west Lake
Turkana, Kenya. Nature, 316, 788-792.
Clark, A.G., Glanowski, S.,
Nielsen, R., Thomas, P.D., Kejariwal, A., Todd, M.A., Tanenbaum, D.M., Civelle,
D., Lu, F., Murphy, B., Ferriera, S., Wang, G., Zheng, X., White, T.J.,
Sninsky, J.J., Adams, M.D., Cargill, M., 2003, Inferring nonneutral evolution
from human-chimp-mouse orthologous gene trios. Science, 302, 1960-1963.
Cantalupo, C., Hopkins, W.D.,
2001, Asymmetric Broca’s area in great apes. Nature, 414, 505.
Deacon, T.W., 1992, The human
brain. Dans The Cambridge Encyclopedia of Human Evolution, édité par S. Jones,
R. Martin et D. Pilbeam (Cambridge: Cambridge University Press), 115-123.
DeGusta, D., Gilbert, W.H.,
Turner, S.P., 1999, Hypoglossal canal size and hominid speech. Proceedings of
the National Academy of Sciences of the USA, 96, 1800-1804.
De Lumley, H., 1998, L’homme
premier. Préhistoire, Evolution, Culture. (Paris: Odile Jacob Ed.), pp. 247.
Falk, D., 1975, Comparative anatomy
of the larynx in man and the chimpanzee: Implications for the language in
Neanderthal. American Journal of Physical Anthropology, 43, 123-132.
Fitch, W. T., Reby, D., 2001,
The descended larynx is not uniquely human. Proceedings of the Royal Society, London,UK, 268, 1669-1675.
Gannon, P.J., Holloway, R.L.,
Broadfield, D.C., Braun, A.R., 1998, Asymmetry of chimpanzee planum temporale:
humanlike pattern of Wernicke’s brain language area homolog. Science, 279,
220-222.
Habib, M., Joanette, Y., Roch
Lecours, A., 2000, Le cerveau humain et les origines du langage.
Médecine/sciences, 16, 171-180.
Heim,
J.-L., 1976, Les Hommes Fossiles de La Ferrassie. Tome I. Le gisement, les
squelettes adultes (crâne et squelette du tronc) (Paris: Archives de L'Institut
de Paléontologie Humaine, mémoire 35, Masson), pp. 397.
Heim, J.-L., Boë, L.-J., Abry,
C., 2002, La parole à la portée du conduit vocal de l’homme de Neandertal.
Nouvelles recherches, nouvelles perspectives. Comptes Rendus Palevol, 1,
129-134.
Heim, J.-L., Boë, L.-J.,
Maeda, S., 2000, Essai de détermination de la position du larynx à partir de
repères craniométriques. Applications à la paléontologie humaine. Dans
L’identité humaine en question. Nouvelles problématiques et nouvelles
technologies en Paléontologie humaine et en Paléoanthropologie biologique,
édité par P. Andrieux, D. Hadjouis et A. Dambricourt-Malassé (Paris: Eds.
Artcom), 187-204.
Holloway, R.L., 1982, Homo
erectus brain
endocasts: volumetric and morphological observations with some comments on cerebral
asymmetries. Actes du Congrès de Nice, Tome 1, 355-369.
Hopkins, W.D., Marino, L.,
Rilling, J.K., MacGregor, L.A., 1998, Planum temporale asymmetries in great
apes as revealed by magnetic resonance imaging (MRI). NeuroReport, 9 (12),
2913-2918.
Jeusel, E., Mafart, B., 2003,
Evolution morphométrique de la cavité palatine des hominidés fossiles.
Anthropologie BRNO,
XLI/3, 205-212.
Johanson, D.C., Taieb M.,
1976, Plio-pleistocene hominid discoveries in Hadar, Ethiopia. Nature, 260,
293-297.
Johanson, D.C., White, T.D.,
1979, A systematic assessment of early African hominids. Science, 203, 321-330.
Jungers, W.L., Pokempner, A.A,
Kay, R.F., Cartmill, M., 2003, Hypoglossal canal size in living hominoids and
the evolution of human speech. Human Biology, 75(4), 473-484.
Kay, R.F., Cartmill, M.,
Balow, M., 1998, The hypoglossal canal and the origin of human vocal behavior.
Proceedings of the National Academy of Sciences of the USA, 95, 5417-5419.
Laitman,
J.T., Crelin, E.S., Colongue, G.J., 1977, The function of the epiglottis in
monkey and man. Yale Journal of Biology and Medecine, 50, 43-48.
Laitman,
J.T., Heimbuch, R.C., 1982, The basicranium of plio-pleistocene hominids as an
indicator of their upper respiratory systems. American Journal of Physical
Anthropology, 59, 323-343.
Laitman, J.T., 1984, The
basicranium and upper respiratory system of African Homo erectus and early Homo sapiens. American Journal of Physical
Anthropology, 63,
180.
Laitman, J.T., 1985, Evolution
of the Hominid Upper Respiratory Tract: The Fossil Evidence. Dans Hominid
Evolution: Past, Present and Future, édité par Tobias Phillp V (New York: Alan
R. Liss, Inc.), 281-286.
Laitman, J.T., 1986, L’origine
du langage articulé. La Recherche, 181, 1164-1173.
Laitman, J.T., Reidenberg,
J.S., Friedland, D.R., Gannon, P.J., 1991, What sayeth thou Neandertal? A look
at the evolution of their vocal tract and speech. American Journal of Physical
Anthropology Suppl., 12, 109.
Laitman,
J.T., Reidenberg, J.S., Marquez, S., Gannon, P.J., 1996, What the nose knows:
New understandings of Neanderthal upper respiratory tract specializations.
Proceedings of the National Academy of Sciences of the USA, 93, 10543-10545.
Leakey,
R. E., 1994, The origin of humankind. (New York: Orion Publishing Group Ltd.
Brockman, Inc.), pp. 192.
Leakey,
L.S.B., 1959, A new fossil skull from Olduvai. Nature, 184, 491-493.
Le Diascorn, H., 1972,
Anatomie et physiologie des sutures de la face. (Paris: Association de la Revue
d’Orthopédie Dento-faciale, Julien Prélat Ed.), pp. 95.
Lieberman, P., 2002, On the
nature and evolution of the neural bases of human language. Yearbook of
Physical Anthropology, 45, 36-62.
Lieberman,
P., 1984, The biology and evolution of language (Cambridge: Harvard University
Press), pp. 379.
Lieberman, P., 1975, On the
origins of language. An introduction to the evolution of human speech (New
York: University Press of America Inc.), pp. 196.
Lieberman, P., 1992, Human
speech and language. Dans The Cambridge Encyclopedia of Human Evolution, édité
par S. Jones, R. Martin, et D. Pilbeam (Cambridge: Cambridge University Press),
134-137.
Marcus, G.F., Fisher, S.E.,
2003, FOXP2 in focus: what can genes tell us about speech and language ?
Trends in Cognitive Sciences, 7(6), 257-262.
Mellars,
P., 1986, A new chronology for the French Mousterian period. Nature, 322,
410-411.
Nishimura, T., Mikami, A.,
Suzuki, J., Matsuzawa, T., 2003, Descent of the larynx in chimpanzee infants.
Proceedings of the National Academy of Sciences of the USA, 100 (12), 6930-6933.
Partridge,
T.C., 1982, The chronological positions of the fossil hominids of Southern
Africa. Dans L'Homo
erectus et la
place de l'Homme de Tautavel parmi les Hominidés fossiles, édité par H. De
Lumley (Nice: 1er
Congrés
International de Paléontologie humaine, prétirage), 617-675.
Paturet, G., 1951, Traité
d’anatomie humaine. Tome I: Ostéologie, Arthrologie, Myologie (Paris: Masson
Ed.), pp. 994.
Pulvermuller, F., 2002, A
brain perspective on language mechanisms: from discrete neuronal ensembles to
serial order. Progress in neurobiology, 67(2), 85-111.
Reeves, H., De Rosnay, J.,
Coppens, Y., et Simonnet, D., 1996, La plus belle histoire du monde. Les
secrets de nos origines. (Paris: Editions du Seuil), pp. 165.
Savage-Rumbaugh, E. S., 1992,
Language training of apes. Dans The Cambridge Encyclopedia of Human Evolution,
édité par S. Jones, R. Martin et D. Pilbeam (Cambridge: Cambridge University
Press), 138-141.
Sherwood, C.C., Broadfield,
D.C., Holloway, R.L., Gannon, P.J., Hof, P.R., 2003, Variability of Broca’s
area homologue in African great apes: Implications for language evolution. The
Anatomical Record Part A, 271A, 276-285.
Swindler, D.R., Wood, C.D.,
1973, An atlas of primate gross anatomy: Baboon, Chimpanzee, and Man
(Washington: University of Washington Press), pp. 370.
Taieb, M., Johanson, D.C.,
Coppens, Y., Aronson, J.L., 1976, Geological and paleontological background of
Hadar hominid site, Afar, Ethiopia. Nature, 260, 289-293.
Takahashi, K., Liu, F.C.,
Hirokawa, K., Takahashi, H., 2003, Expression of Foxp2, a gene involved in
speech and language, in the developing and adult striatum. Journal of
Neuroscience Research, 73(1), 61-72.
Tattersall,
I., 1998,
Becoming Human, Evolution and human uniqueness (New York: Harcourt Brace &
Company), pp.258.
Tobias,
P.V., 1967, The cranium and maxillary dentition of Australopithecus (Zinjanthropus) boisei (Cambridge: Ed. L. S. B.
Leakey, Cambridge University Press), pp. 264.
Valladas,
H. G., Bar-Yosef, O., Vandermeersch, B., 1991, Thermoluminescence dating of
Neanderthal and early modern humans in the Near East. Endeavour, New Series,
15(3), 115-119.
Vandermeersch,
B., 1981, Les hommes fossiles de Qafzeh (Israël) (Paris: CNRS), pp 319.
Vrba,
E., 1982, Biostratigraphy and chronology, based particulary on Bovidae, of southern hominid-associated
assemblages: Makapansgat, Sterkfontein, Taung, Krondraai, Swartkrans; also
Elandsfontein (Saldanha), Broken Hill (now Kabwe) and Cave of Hearth. Dans L'Homo erectus et la place de l'Homme de Tautavel parmi
les Hominidés fossiles, édité par H. De Lumley (Nice: 1er Congrés International de
Paléontologie humaine, prétirage), 707-552.
Walker,
A., Leakey, R., 1993, The skull. Dans The Nariokotome Homo erectus skeleton, édité par A. Walker
et R. Leakey (Berlin: Springer-Verlag), 63-94.
Yokoyama,
Y., Falgueres, C., Lumley, M.-A. de, 1997, Datation directe d'un crâne
proto-Cro-Magnon de Qafzeh par la spectrométrie gamma non destructive. Comptes
Rendus de l’Académie des Sciences série IIa (Paris), 324, 773-779.
Zhang, J., Webb, D. M.,
Podlaha, O., 2002, Accelerated protein evolution and origins of human-specific
features: FOXP2 as an exemple. Genetics, 162, 1825-1835.
Zimmermann, E., 1992, Vocal
communication by non-human primates. Dans The Cambridge Encyclopedia of Human
Evolution, édité par S. Jones, R. Martin et D. Pilbeam (Cambridge: Cambridge
University Press), 124-127.