
Colloques du
Groupement des Anthropologistes de Langue Française (GALF)
Pélissier-Hermitte, B., 2006, Foramens
émissaires et évolution humaine. Antropo, 11, 209-221. www.didac.ehu.es/antropo
Foramens
émissaires et évolution humaine
Emissary
foramens and human evolution
Béatrice Pélissier-Hermitte
UMR 5199, PACEA, Laboratoire d’Anthropologie des
Populations du Passé, Bordeaux 1. Avenue des Facultés 33405 Talence, France.
E-mail: b.pelissier-hermitte@anthropologie.u-bordeaux1.fr
Mots clefs: Drainage émissaire, foramen émissaire, Pléistocène
moyen et supérieur, thermorégulation, évolution humaine.
Key words: Emissary
drainage, emissary foramen, upper middle Pleistocene, thermoregulation, human
evolution.
Résumé
Le drainage veineux cérébral est représentatif de
l’évolution de l’encéphale. Nous étudions une partie de ce drainage, le
drainage émissaire. Afin de l’appréhender, nous examinons des canaux veineux
sur des crânes: les foramens condylaires postérieurs, mastoïdiens, occipitaux,
pariétaux, rétro-articulaires et les foramens nasaux. Nous utilisons deux
approches morphométriques, l’une qualitative (codage), l’autre quantitative
(mesure métrique). L’importance du drainage émissaire semble augmenter avec
l’évolution et la température du milieu.
Abstract
Cerebral venous
drainage is representative of brain evolution. We study a part of this drainage
: the emissary drainage. We examine venous canals on the skull: the posterior
condylar, mastoïd, occipital, parietal, postglenoid and nasal foramina. We use
two morphometrical approaches, the first one, qualitative (scoring) and the
second one, quantitative (measurements). Emissary drainage seems to increase
with evolution and environmental temperature.
Introduction
Pour son fonctionnement et son développement,
l’encéphale a besoin d’une vascularisation qui lui apporte une bonne
oxygénation des tissus et qui contribue à son efficacité et à sa complexité.
Nous étudions une partie du drainage cérébral : le drainage émissaire qui
constitue une voie de suppléance de faible débit. Ce drainage auxiliaire est
composé de différentes veines d’importance variable selon les individus pouvant
même être absentes chez certains. Nous avons donc là des caractères discrets.
Afin d’appréhender la réalité physiologique de ce système veineux auxiliaire,
nous avons examiné des canaux du crâne par lesquels transitent les veines
émissaires homonymes. Il s’agit des foramens émissaires condylaires
postérieurs, mastoïdiens, occipitaux, pariétaux, rétro-articulaires et nasaux,
dont les abréviations sont respectivement: f.C, f.M, f.O, f.P, f.R et f.N.
|
|
1: Veine émissaire
pariétale (parietal
emissary vein); 2: Veine émissaire
occipitale (occipital
emissary vein); 3: Veine émissaire
mastoïdienne (mastoid emissary
vein); 4: Veine émissaire
condylaire (posterior
condyloid emissary vein); 5: Veine émissaire
hypoglosse (hypoglossal
emissary vein); Sa: Sinus sagittal
supérieur (superior
sagittal sinus); R.: Sinus droit (rectus
sinus); T: Sinus transverse
(transverse sinus); Si: Sinus sigmoïde
(sigmoid sinus); O: Sinus occipital (occipital
sinus); M: Sinus marginal (marginal
sinus). D’après Gray (1973) |
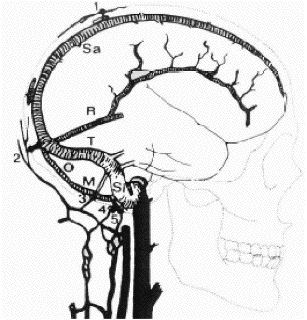
Figure 1.
Veines émissaires et sinus veineux.
Figure 1.
Emissary vein and venous sinuses.
Boyd (1934), Falk (1990), puis Braga (1995) ont réalisé
des études qualitatives de foramens émissaires crâniens principalement sur des
Grands Singes. Braga et Boesch (1997a) ont poursuivi l’étude en augmentant la
qualité de l’échantillon et en y introduisant une approche quantitative. Falk
(1990), Braga et Boesch (1997a,b) ont observé des traces de systèmes veineux
d’hominidés fossiles, essentiellement du Pliocène. Le déterminisme des
caractères discrets est plus ou moins connu et souvent la génétique joue un
rôle considérable, (Crubézy et al., 1999). Rares sont les auteurs ayant étudié les
relations entre l’apparition de ces caractères et les facteurs climatiques.
Silver (1995) ou Hanihara et Ishida (2001) ont examiné les relations entre les
fréquences d’apparition des foramens émissaires et le climat sur des Hommes
actuels.
Dans une optique évolutionniste, Falk (1990) a
soumis l’hypothèse de la «théorie du radiateur» qui repose sur la sensibilité
de l’encéphale à la chaleur et sur des mécanismes mis en oeuvre pour réguler la
température cérébrale. Pendant la réorganisation du réseau veineux cérébral au
cours du temps, certaines veines émissaires auraient vu leur fréquence croître,
elles auraient permis le refroidissement du cerveau dans des conditions
d’hyperthermie. Une meilleure régulation thermique aurait favorisé le
développement et l’augmentation rapide de la taille de l’encéphale dans le
genre Homo. D’après Falk (1990), l’analyse des fréquences d’apparition des
foramens pariétaux et mastoïdiens chez les Grands Singes et les Hommes fossiles
suggère que les veines les plus représentatives du développement du cerveau
sont les veines pariétales et mastoïdiennes. De plus, Cabanac et Brinnel (1985)
remarquent l’importance de ces deux veines émissaires dans la thermorégulation.
Braga et Boesch (1997b) ne partagent pas cette idée et pensent que les foramens
mastoïdiens ne représentent pas des caractères dérivés, car ils ont une
fréquence d’apparition assez élevée chez les Grands Singes (71,4% chez les Pan
troglodytes et 89,5% chez les Pan paniscus) et ont aussi une grande
variabilité de fréquences. Il existe une controverse sur la théorie du
radiateur. Holloway (1990) remarque qu’il n’existe pas de preuve que des
contraintes thermiques influencent le cerveau et ajoute qu’il est difficile de
savoir si l’augmentation du nombre de veines pariétales est due au
refroidissement du cerveau. Différentes causes physiologiques peuvent être à
l’origine d’un tel phénomène. Scheibel (1990) s’interroge aussi sur
l’importance du drainage des veines émissaires pour opérer une diminution de la
température du cerveau.
Cabanac (1986) ainsi que Zenker et Kubik (1996)
notent que, dans des conditions d’hyperthermie, le cerveau se refroidit grâce à
une réorganisation du système veineux. Falk (1990) évoque le fait que certaines
fréquences de foramens émissaires augmentent dans un milieu chaud. Ces diverses
études invitent à se poser davantage de questions sur les relations entre la
vascularisation du drainage émissaire et l’environnement.
A l’aide de mesures qualitatives et quantitatives qui
permettent d’estimer l’importance des drainages émissaires sur des échantillons
d’Hommes pléistocènes et actuels, deux aspects évolutifs sont abordés et
présentent de nombreuses interconnections : la macro et la microévolution. La
macroévolution représente ici l’évolution des hominidés du Pliocène jusqu'à la
fin du Pléistocène. La microévolution a un cadre chronologique plus limité et
concerne, dans l’étude, les phénomènes d’adaptations climatiques,
particulièrement, l’adaptation au froid des Néandertaliens et des Groenlandais.
L’influence du climat sur le drainage émissaire a été peu étudiée chez les
Hommes actuels, mais c’est la première fois que l’on examine ces relations sur
des sujets de la lignée néandertalienne.
Dans une optique microévolutive, j’ai comparé dans
un premier temps deux échantillons de populations d’Hommes actuels vivant dans
des milieux où la température diffère nettement: Groenland et Gabon. Dans un
second temps, j’ai effectué une comparaison de foramens émissaires chez des
Hommes de lignée néandertalienne. Nous portons notre intérêt sur le facteur
thermique qui n’est qu’un des multiples paramètres du milieu. Les résultats
seront donc à prendre avec prudence.
Matériel
J’ai examiné deux échantillons d’Hommes actuels, (30
sujets du Gabon et 37 du Groenland [Musée de l’Homme de Paris]) et quelques
Hommes du Pléistocène moyen et supérieur (moulages de l’Université de Bordeaux
1): Pré-néandertaliens (N = 2), Néandertaliens (N = 20) et Hommes
anatomiquement modernes (N = 19). J’ai aussi utilisé des informations issues de
la littérature relatives à des hominidés. Cf. Tab.1.
Rappelons que certains fossiles, par exemple, Gran Dolina (Atapuerca, Espagne)
font l’objet d’un débat sur leur systématique, (Arsuaga et al., 2001, Bermudez et
al., 2004). Certains fossiles sont parfois attribués à Homo antecessor (crâne 5
d’Atapuerca), (Bermudez de Castro et al., 1997) ou à Homo heidelbergensis
(Pétralona),
(Grün, 1996). Ils sont aussi souvent considérés par d’autres auteurs comme des
Pré-néandertaliens. Je classe tous ces fossiles au sein des Pré-néandertaliens,
sachant qu’une étude plus approfondie pourrait se révéler instructive.
Les hominidés connaissent une vaste répartition
chronologique et géographique. Les Néandertaliens d’Europe centrale de Krapina
habitaient dans un milieu froid (130 kA) et Amud 1 (53 kA), par exemple, vivait
dans le milieu plus chaud du Proche-Orient, (Minugh-Purvis et al., 2000 et Rink et
al., 2001).
Dans un premier temps, nous considérons que les
Pré-néandertaliens et les Néandertaliens d’Europe vivaient dans un milieu froid
et les Néandertaliens du Proche-Orient dans un milieu chaud. Cette première
approche a pour but de dégager d’éventuelles grandes lignes directrices. Dans
un second temps, afin de préciser l’étude, nous séparerons les sujets de la
lignée néandertalienne selon leur stade isotopique. Nous ne discriminerons pas
les Hommes anatomiquement modernes de cette manière, car ils ont presque tous
vécu au cours du stade isotopique 2. Cf. Tab. 1.
|
Noms / Names |
St. isotop. |
Noms / Names |
St. Isotop. |
|
a. Homo sapiens anatomiquement
moderne |
|||
|
Barahonda |
|
Galley Hill * |
|
|
Brno II |
3/2 *4 |
Mechta et Arbi * |
2 *22 |
|
Chancelade 1 |
2 |
Taforalt * |
2 *19,22 |
|
Cioclovina 1 |
3/2 |
Kanjera 1 * |
|
|
Cro-magnon 1 |
2 *4 |
Kanjera 3 * |
|
|
Cro-magnon 2 |
2 *4 |
Kelsterbach * |
3/2 |
|
Dolni
Vestonice 2 |
2
*3 |
KNM-ER
5306 * |
2/1
*22 |
|
Fallah
H16 |
2 |
KNM-ER
5310 * |
2/1
*22 |
|
Grotte
des enfants 6 |
2 |
KNM-ER
5493g * |
2/1
*22 |
|
Janislawice |
2/1 |
KNM-ER 5525 * |
2/1 *22 |
|
Lagoa Santa |
1 |
KNM-ER 7459 * |
2/1 *22 |
|
Minatogawa 1 |
2 *1 |
KNM-ER 7461a * |
2/1 *22 |
|
Minatogawa 2 |
2 *1 |
KNM-ER 7465 * |
2/1 *22 |
|
Minatogawa 4 |
2 *1 |
KNM-ER 7480b * |
2/1 *22 |
|
Mumba X |
2 |
KNM-ER 8524e * |
2/1 *22 |
|
Mladec V |
3/2 *2 |
KNM-ER 8424f * |
2/1 *22 |
|
Mladec VI |
3/2 *2 |
H. des Cottés* |
3/2 *22 |
|
Pavlov |
1 2 *2 |
M 17017 * |
2
*20 |
|
Starselje |
3/2 |
Predmost III * |
? *21 |
|
Camargo * |
3/2 |
Predmost IV * |
? *21 |
|
Cro-magnon III * |
2 *17 |
Predmost IX * |
? *21 |
|
Dolni-Vestonice III* |
2 *22 |
Predmost X * |
? *21 |
|
EM 1326 * |
2*20 |
Vogelherd 1 * |
3/2 *18 |
|
EM 523 * |
2*20 |
Border Cave * |
|
|
EM 525 * |
2*20 |
Brno III * |
3/2 *21 |
|
EM
550 * |
2*20 |
Kow Swamp 7 * |
2/1 *21 |
|
b. Néandertaliens du Proche-Orient |
|||
|
Amud 1 |
3 *12 |
Tabun 1 |
3/2 *16 |
|
Qafzeh 3 |
5 *13 |
Shanidar 5 |
3 *14 |
|
Qafzeh 9 |
5 *13 |
Teschik-Tasch |
4 *2 |
|
Qafzeh 11 |
5 *13 |
|
|
|
c. Néandertaliens d’Europe |
|||
|
Chap.
aux Saints |
3
*2 |
Gibraltar
II * |
3
*21 |
|
Circeo
1 |
3
*7,8 |
Marillac
n°72 * |
3
/ 4 *23 |
|
Engis
1 |
3 |
Moustier
* |
3
/4 *25 |
|
Ferrassie |
3/2
*3,6 |
Hortus
XLIX * |
2*23 |
|
Fontéchevade |
6
*4 |
Krapina
10 * |
6/5*24 |
|
Gibraltar
1 |
3,4
*17 |
Krapina
2 * |
6/5*24 |
|
Krapina
1 |
5
*10,11 |
Krapina
34 17 * |
6/5*24 |
|
Krapina
3 |
5
*10,11 |
Krapina
43 * |
6/5*24 |
|
La
Quina H5 |
3
*4 |
Krapina
6 * |
6/5*24 |
|
Néandertal |
4/3
*9 |
Krapina
A * |
6/5*24 |
|
Saint
Césaire |
3/2
*2 |
Neanderthal
* |
3
/4 *25 |
|
Spy
1 |
3
*5 |
Saccopastore
* |
5
*26 |
|
Spy
2 |
3
*5 |
|
|
*1:
Kamminga et Wright (1988); *2: www.
archaeologyinfo.com; *3: Trinkaus et al. (2000);
*4: Saban (1984); *5: www.modernhumanorigins.com; *6:
Leroyer (1988); *7: Manzi et al (1996);
*8: Coutselinis et al. (1991); *9:
Dean et al. (1998); *10: Alemseged et
al. (2002); *11: Minugh-Purvis et
al. (2000); *12: Rink et al.
(2001);*13: Schwarcz et
al. (1988); *14: Anton (1997); *15:
Grün (1996); *16: Schwarcz et al. (1998);
*17: Oakley (1964); *17:
Henry-Gambier (2002); *18: Conard et al. (2004); *19:
Bonfiglioli et al. (2004); *20:
Falk (1986); * 21: Arsuaga et al. (1999);
*22: Saban (1984); *23:
www. archaeologyinfo.com; *24: Alemseged et al. (2002);
*25: Dean et al. (1998); *26:
Manzi et al. (1996); *27:
modernhumanorigins.com; *28: Bermudez et al. (2004);
*29: Falguères et al. (1999);
SH: Sima de los Huesos; GD.:
Gran Dolina.
Tableau 1.
Échantillon d’Hommes du Pléistocène moyen et supérieur. Les sujets dont le nom
est suivi d’un astérisque viennent de la littérature.
Table 1.
Upper and Middle Pleistocene hominids’ samples. Fossil’s name which followed by
an asterisk come from literature.
|
Noms / Names |
St. isotop. |
Noms / Names |
St. Isotop. |
|
d. Pré-néandertaliens |
|||
|
Biache 1 |
6 *4 |
AT 928 (SH) * |
7 -12*28 |
|
Pétralona 1 |
6,7 *8,15 |
ATD 16 (GD) * |
< 18 *29 |
|
Arago
21 * |
7
-10*27 |
ATD
57 (GD) * |
<
18 *29 |
|
AT-1122
(SH) * |
7-12*28 |
ATD
16 (GD) * |
<
18 *29 |
|
AT
125 (SH) * |
7-12*28 |
ATD
57 (GD) * |
<
18 *29 |
|
AT
1661 (SH) * |
7-12*28 |
cranium
4 (GD) * |
<
18 *29 |
|
AT
365 (SH) * |
7-12*28 |
cranium
5 (GD) * |
<
18 *29 |
|
AT
395 (SH) * |
7-12*28 |
cranium
6 (GD) * |
<
18 *29 |
|
AT
396 (SH) * |
7-12*28 |
cranium
7 (GD) * |
<
18 *29 |
|
AT
425 (SH) * |
7-12*28 |
H.
de la Chaise * |
6
*22 |
|
AT
644 (SH) * |
7-12*28 |
Cova
Negra * |
6
*22 |
|
AT
84 (SH) * |
7-12*28 |
|
|
Tableau 1.
Cont.
Table
1. Cont.
Méthodes
Deux types de mesures ont été effectués: des mesures
qualitatives et quantitatives. L’interprétation des résultats qualitatifs
permet de classer des fréquences d’apparition en deux catégories. La première,
individuelle, correspond à la présence du caractère sur au moins un des côtés:
[(Nd ou Ng)/Nind] x 100; la seconde, par côté: [(Nd et Ng)/(Nind x 2)] x 100.
Des mesures quantitatives permettent d’estimer le
drainage moyen passant par les veines émissaires grâce à la surface de section
elliptique des foramens émissaires. A l’aide d’un pied à coulisse électronique
de précision 0,02 mm, nous mesurons les deux rayons de cette surface et en
estimons l’aire en mm2, (Pélissier-Hermitte, nd).
Lorsque les différents échantillons sont comparés
selon l’angle qualitatif, nous employons les tests de Chi2 ou de Fischer. Quand
il s’agit d’une comparaison des résultats quantitatifs des divers échantillons,
nous utilisons le test de Mann et Whitney.
Résultats
Observations sur l’ensemble des échantillons
Les tableaux 2 et 3 rassemblent les résultats
observés sur l’ensemble des échantillons.
Les fréquences d’apparition des foramens émissaires
(individuelles et par côté) sont toujours supérieures chez les Hommes actuels
par rapport aux Hommes fossiles, à l’exception des foramens rétro-articulaires
des Néandertaliens. Très souvent, que ce soit chez les Néandertaliens ou chez
les Hommes actuels, les fréquences d’apparition sont plus conséquentes dans les
échantillons vivant dans un environnement chaud. Souvent, les drainages
émissaires sont plus élevés chez les sujets vivant dans les milieux les plus
chauds. Cf. Tab. 2 et 3.
Symétrie-Asymétrie
Le tableau 4 permet de comparer sur des échantillons
les fréquences d’apparition individuelles et par côté.
Tous les foramens sont plus nombreux unilatéralement
que bilatéralement.
Étude multivariée des fréquences d’apparition -
Analyse en composantes multiples: ACM.
Chez les Hommes pléistocènes, des données sont
souvent manquantes (partie osseuse absente). Ces informations manquantes n’ont
aucune valeur biologique et les intégrer dans l’ACM pourrait biaiser les
résultats. Nous avons supprimé des individus et les informations relatives aux
foramens condylaires et nasaux qui sont souvent absents chez les Hommes
fossiles. Les fossiles exclus sont Minatogawa 4, Mumba X, Pavlov 1, Dolni
Vestonice 2, Mladec V et VI, Brno 2, Cro-magnon 2, Janislawice, Engis 1, la
Quina H5, Gibraltar 1, Fontéchevade 5, Qafzeh 3 et 9, Shanidar 5, et Pétralona.
L’ACM est composée de 20 variables: fMd, fMg
(foramens mastoïdiens droits et gauches), fOd, fOg (foramens occipitaux droits
et gauches), fPd, fPg (foramens pariétaux droits et gauches), et fRd, fRg
(foramens rétro-articulaires droits et gauches). Un foramen peut être classé en
trois catégories: absent, unique ou multiple. Dans le dernier cas, propre aux
foramens mastoïdiens et occipitaux, les ouvertures peuvent être doubles ou
triples. Les foramens rétro-articulaires ne sont jamais multiples. Les
individus sont associés à l’ACM comme variables illustratives, leur importance
n’influe pas sur la variance.
Le figure 2 représente 26,42% de la variance totale.
Les foramens multiples sont localisés dans la partie droite du graphique. Ce
sont des foramens rares, mais leur présence influence notablement l’analyse.
Les foramens uniques sont situés à proximité des Hommes actuels alors que les
foramens absents sont regroupés en général près des Hommes pléistocènes. Les
sujets vivant sous les climats les plus chauds ont davantage de foramens
émissaires. Les Pré-Néandertaliens sont plus proches des Néandertaliens
d’Europe que de ceux du Proche-Orient. Il semble que le nombre de foramens augmente
avec le temps et la température du milieu.
|
|
|
H. fossiles / fossil
H. |
H. actuels / actual
H. |
|||||
|
|
|
PN |
NE |
Npo |
H.anat. |
H. act. |
Groenl |
Gabon |
|
f.C |
Fréq. Ind |
0,00 |
0,00 |
14,29 |
10,91 |
83,14 |
94,59 |
76,67 |
|
Fréq. Côté |
0,00 |
0,00 |
7,14 |
6,36 |
69,14 |
87,84 |
66,67 |
|
|
f.M |
Fréq. Ind |
45,80 |
19,35 |
57,14 |
36,36 |
89,14 |
78,38 |
93,33 |
|
Fréq. Côté |
25,00 |
9,68 |
28,57 |
26,36 |
75,71 |
63,51 |
73,33 |
|
|
f.O |
Fréq. Ind |
4,17 |
9,68 |
28,57 |
0,00 |
73,43 |
40,54 |
80,00 |
|
Fréq. Côté |
2,08 |
6,45 |
14,29 |
0,00 |
55,71 |
24,32 |
65,00 |
|
|
f.P |
Fréq. Ind |
4,17 |
6,45 |
14,29 |
16,36 |
67,71 |
72,97 |
73,33 |
|
Fréq. Côté |
2,08 |
4,84 |
7,14 |
12,73 |
50,86 |
54,05 |
56,67 |
|
|
f.R |
Fréq. Ind |
0,00 |
9,68 |
28,57 |
0,00 |
14,57 |
8,11 |
23,33 |
|
Fréq. Côté |
0,00 |
4,84 |
21,43 |
0,00 |
8,71 |
4,05 |
13,33 |
|
|
f.N |
Fréq. Ind |
4,17 |
6,45 |
28,57 |
5,45 |
70,29 |
54,05 |
73,33 |
|
Fréq. Côté |
2,08 |
4,84 |
14,29 |
2,73 |
58,14 |
40,54 |
58,33 |
|
H. act: Échantillon
regroupant les sujets du Groenland et du Gabon (Sample of individuals’
Greeland and Gabon) ; PN: Pré-Néandertaliens (Pre-neandertals)
; NE: Néandertaliens d’Europe (European Neandertals)
; Npo: Néandertaliens du Proche-Orient (Levantine Neandertals)
; H. anat.: Hommes anatomiquement modernes (morphological modern hominids).
Tableau 2.
Fréquences relatives d’apparition individuelles et par côté (%) . Les mesures
qualitatives sont des fréquences d’apparition exprimées en %. Des données de la
littérature sont ajoutées aux données observées. Lorsque le caractère est
absent, non explicitement décrit dans la littérature ou lorsque la partie
osseuse sur laquelle il pourrait éventuellement être présent, est cassée, le
codage est de 0.
Table 2.
Relative individual frequency of apparition and relative frequency by side
(%).Frequencies of apparition are qualitative measurements. In qualitative
study, we used literature’s data and observed data. The scoring is 0, when the
character is absent, not explicitly describe in the literature, or when the
osseous remain is broken.
|
|
H. fossiles / fossil
H. |
H. H. actuels /
actual H. |
|||||
|
|
PN |
NE |
Npo |
H.anat. |
H. act. |
Groenl |
Gabon |
|
f.C |
0,00 |
0,00 |
4,46 |
6,128 |
6,67 |
5,37 |
7,16 |
|
f.M |
4,45 |
13,45 |
5,51 |
6,68 |
3,85 |
3,24 |
3,73 |
|
f.O |
1,64 |
1,55 |
1,09 |
5,97 |
1,96 |
1,61 |
1,92 |
|
f.P |
0,00 |
0,00 |
2,92 |
2,64 |
1,92 |
1,61 |
1,86 |
|
f.R |
0,00 |
0,83 |
2,36 |
2,31 |
0,88 |
1,02 |
1,33 |
|
f.N |
3,07 |
0,00 |
1,91 |
0,94 |
0,85 |
0,79 |
1,02 |
H. act: Échantillon
regroupant les sujets examinés du Groenland et du Gabon (Sample Greeland and
Gabon); PN: Pré-néandertaliens (Pre-neandertals);
NE: Néandertaliens (European Neandertals);
Npo: Néandertaliens du Proche-Orient (Levantine Neandertals)
Hommes anatomiquement modernes (morphological modern hominids).
Tableau 3.
Surfaces de section des foramens émissaires. Les mesures quantitatives sont les
moyennes des surfaces des foramens (mm2).
Table 3.
Area’s section of the emissary foramina. Average areas are quantitative
measures (ellipse, mm2).
|
|
H. fossiles / fossil
H. |
H. actuels / actual
H. |
|||||
|
|
PN |
NE |
Npo |
H.anat. |
H. act. |
Groenl |
Gabon |
|
f.C |
1,0000 |
1,0000 |
0,0231 |
0,3079 |
0,0346 |
0,2643 |
0,3318 |
|
f.M |
0,0783 |
0,1930 |
0,2191 |
0,1869 |
0,0254 |
0,1149 |
0,0279 |
|
f.O |
0,6125 |
0,5792 |
0,4418 |
1,0000 |
0,0158 |
0,0808 |
0,1468 |
|
f.P |
0,6125 |
0,7463 |
0,0231 |
0,5266 |
0,0242 |
0,0573 |
0,1279 |
|
f.R |
1,0000 |
0,3729 |
0,7212 |
1,0000 |
0,2060 |
0,3744 |
0,2333 |
|
f.N |
0,6125 |
0,7463 |
0,4418 |
0,3802 |
0,0956 |
0,1803 |
0,1674 |
Tableau 4.
Degré de significativité entre les fréquences d’apparition individuelles et par
côté.
Table 4.
Degree of significance between individual and by side frequencies of apparition
(%).
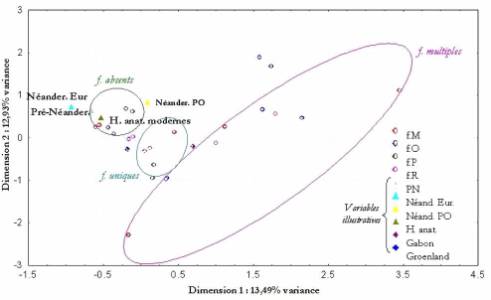
Figure 2. ACM. Types de
foramens émissaires – échantillon d'hominidés.
Figure 2. ACM. Emissary
foramina' types – hominids' samples.
Différences de fréquences et de surfaces entre les différents échantillons
Le tableau 5 indique la significativité des
différences quantitatives et qualitatives observées entre les différents
échantillons.
|
|
|
p.
H.act-H.foss |
p
N-Groen. |
P
N-Gab |
p
N-H.anat. |
p.
Ne-Npo |
p.
PN-Npo |
p
Gab-Croen |
|
f.C |
Fréq.
Ind |
0,3514 |
0,4800 |
0,3529 |
0,2350 |
0,1842 |
0,2258 |
0,0671 |
|
|
Fréq.
Côté |
0,3071 |
0,4400 |
0,3015 |
0,1442 |
0,1842 |
1,0000 |
0,7836 |
|
|
Surf.
/ Area |
0,0065* |
0,0240* |
0,0336* |
0,6434 |
1,0000 |
1,0000 |
0,2780 |
|
f.M |
Fréq.
Ind |
0,5297 |
0,5200 |
0,5588 |
0,3191 |
0,0625 |
0,6851 |
0,1659 |
|
|
Fréq.
Côté |
0,3859 |
0,3800 |
0,3971 |
0,2097 |
0,0799 |
0,1463 |
0,6791 |
|
|
Surf.
/ Area |
0,0366* |
0,4900 |
0,1779 |
0,6797 |
0,5637 |
0,3545 |
0,2269 |
|
f.O |
Fréq.
Ind |
0,2541 |
0,2667 |
0,4265 |
0,1145 |
0,2226 |
0,1199 |
0,5821 |
|
|
Fréq.
Côté |
0,1793 |
0,1600 |
0,3309 |
0,0649 |
0,3038 |
0,2466 |
0,4254 |
|
|
Surf.
/ Area |
0,0174* |
0,0209* |
0,0025* |
0,0641 |
0,1213 |
1,0000 |
0,1306 |
|
f.P |
Fréq.
Ind |
0,3351 |
0,4000 |
0,3676 |
0,3490 |
0,4630 |
0,4065 |
0,7313 |
|
|
Fréq.
Côté |
0,2527 |
0,2933 |
0,2794 |
0,0968 |
0,5468 |
0,6478 |
0,5522 |
|
|
Surf.
/ Area |
0,2076 |
0,5940 |
0,1939 |
0,8273 |
1,0000 |
1,0000 |
0,1367 |
|
f.R |
Fréq.
Ind |
0,0811 |
0,7110 |
0,1765 |
0,0091* |
0,2226 |
0,0452* |
0,0982 |
|
|
Fréq.
Côté |
0,0462* |
0,4946 |
0,1029 |
0,0041* |
0,0722 |
0,0808 |
0,0630 |
|
|
Surf.
/ Area |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
|
f.N |
Fréq.
Ind |
0,2703 |
0,3200 |
0,3824 |
0,4349 |
0,1475 |
0,2207 |
0,6269 |
|
|
Fréq.
Côté |
0,2011 |
0,2333 |
0,2941 |
0,2749 |
0,2269 |
0,4036 |
0,4851 |
|
|
Surf.
/ Area |
0,8495 |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
1,0000 |
0,0387* |
N:
Néandertaliens d’Europe et du Proche-Orient (European and levantine
Neandertals);
Fréq.
ind.: individual frequency ; Fréq. côté: by side frequency
Tableau
5. Significativité des différences entre
échantillons. Les tests de Chi2 ou de Fischer (italique) sont
bilatéraux. Le seuil d’acceptation de l’hypothèse est de 5%. Les probabilités
significatives sont suivies d’un astérisque.
Table
5. Significantly between samples’ differences. Chi2
test and Fischer test (in italics) are on the both sides. Hypothesis threshold
is 5%. Significant probabilities are followed by an asterisk.
La lignée néandertalienne
Le tableau 6 regroupe les résultats observés sur les
échantillons de la lignée néandertalienne et classés selon la température du
milieu.
|
|
|
Pré-néandertalien
/ Pre-neandertals |
Néandertalien
/ Neandertalien |
||||||
|
|
|
|
|
|
Europe |
Poche
Orient/ Near
East |
|||
|
Temperature |
Froid
/ Cold |
Tempéré
/ Temperate |
Chaud
/ Hot |
Froid
/ Cold |
Tempéré
/ Temperate |
Chaud
/ Hot |
Tempéré
/ Temperate |
Chaud
/ Hot |
|
|
Stade
isotop. |
6 |
12
; 7 *2 |
<18
*1 |
6
;2 |
4
;3 |
5 |
>
5 *3 |
5
*3 |
|
|
f.C |
Fréq.
Ind |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
25,00 |
0,00 |
|
|
Fréq.
Côté |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
12,50 |
0,00 |
|
|
Surf.
/ Area |
0,00 |
- |
- |
0,00 |
0,00 |
0,00 |
1,11 |
0,00 |
|
f.M |
Fréq.
Ind |
50,00 |
26,26 |
50,00 |
33,33 |
44,44 |
40,00 |
75,00 |
33,33 |
|
|
Fréq.
Côté |
50,00 |
13,33 |
25,00 |
16,66 |
27,77 |
20,00 |
37,50 |
16,66 |
|
|
Surf.
/ Area |
6,19 |
- |
- |
0,00 |
1,94 |
4,16 |
4,80 |
0,95 |
|
f.O |
Fréq.
Ind |
50,00 |
6,66 |
0,00 |
0,00 |
22,22 |
0,00 |
25,00 |
33,33 |
|
|
Fréq.
Côté |
25,00 |
3,33 |
0,00 |
0,00 |
11,11 |
0,00 |
12,50 |
16,66 |
|
|
Surf.
/ Area |
1,64 |
- |
- |
0,00 |
0,39 |
0,00 |
0,29 |
0,34 |
|
f.P |
Fréq.
Ind |
0,00 |
0,00 |
0,00 |
0,00 |
10,00 |
0,00 |
50,00 |
0,00 |
|
|
Fréq.
Côté |
0,00 |
0,00 |
0,00 |
0,00 |
5,00 |
0,00 |
37,50 |
0,00 |
|
|
Surf.
/ Area |
0,00 |
- |
- |
0,00 |
0,00 |
0,00 |
1,46 |
0,00 |
|
f.R |
Fréq.
Ind |
0,00 |
0,00 |
0,00 |
33,33 |
11,11 |
10,00 |
25,00 |
33,33 |
|
|
Fréq.
Côté |
0,00 |
0,00 |
0,00 |
16,66 |
5,55 |
5,00 |
25,00 |
16,66 |
|
|
Surf.
/ Area |
0,00 |
- |
- |
0,26 |
0,11 |
0,00 |
1,01 |
0,23 |
|
f.N |
Fréq.
Ind |
0,00 |
0,00 |
0,00 |
0,00 |
11,11 |
10,00 |
25,00 |
0,00 |
|
|
Fréq.
Côté |
0,00 |
0,00 |
0,00 |
0,00 |
5,55 |
10,00 |
12,50 |
0,00 |
|
|
Surf.
/ Area |
0,00 |
- |
- |
0,00 |
0,00 |
0,00 |
0,48 |
0,00 |
*1:
Bermudez et al. (2004); *2: Iacumin et al. (1996);
*3: Shea (2003)
Tableau
6. Fréquences relatives d’apparition individuelle et
par côté (%) ainsi que la surface de section des foramens émissaires (mm2) de
Néandertaliens. Seuls les sujets dont j’ai la datation sont inclus dans le
tableau 6. Cf. Tab 1. Fréq. Ind. : Fréquence individuelle ; Fréq.
côté : Fréquence par côté.
Table
6. Relative individual frequencies of apparition and
relative frequencies by side (%), and area’s section of the emissary foramina
(mm2) of Neandertals. Only fossils who have dating are presents in table 6.
(Tab. 1). Fréq. Ind. :individual frequency ; Fréq. côté : by side frequency.
Seul, dans tout l’échantillon d’hominidés de la
lignée néandertalienne, Teschik-Tasch possède un foramen condylaire (droit). On
observe une absence de ces foramens sur la Ferrassie, Spy 1, les crânes 5 et 6
d’Atapuerca et Qafzeh 11. Dans 25 cas sur 51 (49%) cette partie de l’occipital
est cassée. Dans les autres cas, je n’ai pas trouvé d’informations relatives à
ce foramen dans la littérature.
Aucun foramen nasal n’est présent dans l’échantillon
de Pré-néandertaliens. La majorité des informations vient de crânes d’Atapuerca
(Espagne) où de nombreux restes sont
fragmentaires. Sur les calvaria presque complètes d’Atapuerca, on
remarque une absence de foramen pour les crânes 4 et 5, et des os nasaux cassés
sur les crânes 6 et 7. Cova Négra, Fontéchevade et Biache Saint Vaast ont aussi
des os nasaux cassés, ceux d’Arago 21 sont préservés et montrent une absence de
foramen. Pétralona 1 présente deux foramens nasaux à chacun des deux côtés
(gauche et droit). Ce Pré-néandertalien est exclu de l’étude, car il vivait au
stade isotopique 6 ou 7. Or, le stade isotopique 6 est une période glacière et
pendant le stade 7 régnait un milieu tempéré froid, Cf. Tab. 1. Les
foramens pariétaux et rétro-articulaires sont assez rares dans la lignée
néandertalienne. Cf. Tab. 6.
Les foramens apparaissent généralement
unilatéralement. Seuls les foramens rétro-articulaires des Néandertaliens les
plus tardifs du Proche-Orient et les foramens mastoïdiens des
Pré-néandertaliens du stade 6 se révèlent être uniquement des foramens
bilatéraux. Biache 1 a des foramens mastoïdiens symétriques et Teschik-Tasch
possède des foramens rétro-articulaires à droite et à gauche. Cf. Tab. 5 et 6
Étude multivariée des fréquences d’apparition: ACM
Comme dans la première analyse en composantes
multiples, nous n’avons pas tenu compte des informations relatives aux foramens
condylaires et nasaux, nous excluons aussi les foramens rétro-articulaires.
L’analyse repose sur 16 hominidés. L’ACM est composée de 8 variables: trois
types de foramens émissaires sont présents, les mastoïdiens, occipitaux et
pariétaux. Chaque foramen a trois modalités d’apparition: absente (a), unique
(u) ou multiple (m). Les individus sont associés à l’ACM comme variables
illustratives. Le graphique représente 57,36% de la variance totale.
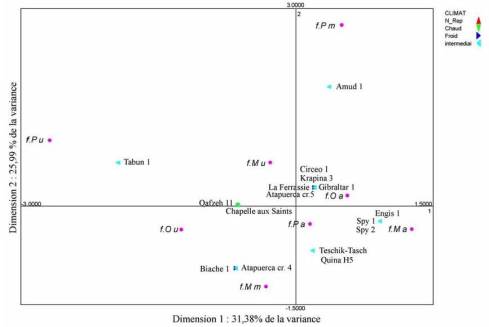
Figure 3.
ACM . Types de foramens émissaires – lignée néandertalienne
Figure 3.
ACM . Emissary foramina’s types – Neandertalian lineage
La majorité des fossiles, surtout des Néandertaliens
européens est regroupée en bas à droite du graphique, là où sont localisées les
variables représentant les foramens absents. Un important groupe de ces
Néandertaliens est situé à proximité de la variable indiquant la présence d’un
foramen mastoïdien unique. C’est ce type de foramen émissaire qui est le plus
fréquemment observé chez ces hominidés. Cf. Tab. 2 et 6. Les foramens
occipitaux ne présentent pas de foramens multiples. Tabun 1 se distingue des
autres car il a des foramens mastoïdiens gauches doubles ainsi que deux
foramens rétroarticulaires. L’autre fossile atypique, qui est aussi un Néandertalien
du Proche-Orient, est Amud 1. Il est le seul hominidé à présenter un foramen
pariétal. Les Prénéandertaliens de Biache 1 et du crâne 4 d’Atapuerca ont la
particularité d’avoir des foramens mastoïdiens multiples et un foramen
occipital simple. Le crâne 5 d’Atapuerca est plus proche des Néandertaliens
d’Europe avec seulement un foramen mastoïdien simple.
Discussion
Macroévolution
Les résultats qualitatifs et quantitatifs des Hommes
actuels sont tous supérieurs à ceux des Hommes pléistocènes. C’est entre les
deux échantillons d’Hommes actuels et d’Hommes pléistocènes que s’observe le
plus grand nombre de différences, particulièrement en ce qui concerne les
résultats quantitatifs. (Le nombre important de sujets des deux échantillons
doit influencer les résultats des tests statistiques.)
D’un point de vue quantitatif, le drainage des
foramens émissaires condylaires, mastoïdiens et occipitaux augmente avec le
temps. Nous observons avec ce même paramètre une augmentation significative de
la fréquence des foramens rétroarticulaires, Cf. Tab. 5. Or, ce
caractère est souvent considéré comme un caractère primitif, (Saban, 1984).
Aucun foramen rétro-articulaire n’est observé parmi les échantillons de
Pré-néandertaliens et d’Hommes anatomiquement modernes. Quelques uns existent
chez les Néandertaliens, particulièrement chez ceux du Proche-Orient; 3
foramens rétro-articulaires pour six individus, sachant que Teschik-Tasch en
comprend deux. Cf. Tab. 2, 3 et 5.
Macroévolution - Climat
La fréquence individuelle des foramens
rétroarticulaires diffère entre les Pré-néandertaliens et les Néandertaliens du
Proche-Orient. Les différences significatives entre, d’une part les
Néandertaliens et les Groenlandais et, d’autre part, entre les Néandertaliens
et les Gabonais sont similaires. La différence majeure semble plutôt provenir
de la différence entre Néandertaliens et Hommes actuels plutôt qu’entre les
différences de milieux. Cf. Tab. 5
Microévolution - Climat
Les différences qualitatives entre les échantillons
de Néandertaliens d’Europe et du Proche-Orient ne sont jamais significatives.
Notons, toutefois, que les fréquences d’apparition individuelles et par côté
des foramens mastoïdiens des Néandertaliens du Proche-Orient sont presque
supérieures à celles des Néandertaliens d’Europe. Seule la surface du foramen
nasal diffère entre les sujets du Gabon et du Groenland. Cf. Tab. 2, 3 et 5.
Analyses multivariées
La première ACM (figure 2) paraît confirmer les autres observations
qualitatives en indiquant que le nombre de foramens mastoïdiens, occipitaux,
pariétaux et rétroarticulaires augmente avec le temps et la température.
Lorsque nous étudions uniquement la lignée néandertalienne sur le figure 3,
nous observons globalement que les Néandertaliens ont peu de foramens
émissaires et qu’il s’agit principalement de foramens mastoïdiens. Les
Néandertaliens du Proche-Orient paraissent avoir davantage de foramens
émissaires que ceux d’Europe, mais aucun test ne montre de différences
significatives. Cf. Tab. 5
Foramens mastoïdiens
Maintenant, nous observons plus en détail les
foramens mastoïdiens, les plus nombreux parmi les foramens présents sur les
Hommes fossiles. Cf. Tab. 7
|
|
H. actuels/ Actual H. N = 67 |
H. fossiles/ Fossil
H. N = 41 |
|
f. absents / absent
f. |
42 (31,34%) |
55 (67,07%) |
|
f. simples / single
f. |
63 (47,01%) |
20 (24,39) |
|
f. doubles /double
f. |
25 (18,66%) |
7 (8,54%) |
|
f. triples / triple
f. |
4 (2,99%) |
0 (0,00%) |
Tableau 7.
Nombre et fréquence par côté (%) de foramens mastoïdiens des échantillons.
Table 7.
Number and by sides frequencies (%) of mastoid foramina in samples.
L’absence de foramen mastoïdien dans l’échantillon
d’Hommes pléistocènes est le cas le plus fréquent, comme le montre le degré de
significativité de 0,0002. Les Hommes pléistocènes ont moins de foramens à
ouverture simple ou multiple que les Hommes actuels. Les différences ne sont
pas significatives, même si le degré de significativité des foramens simples
approche du seuil de 5%: p = 0,0645. Ces résultats rejoignent ceux de la
première ACM (Cf. Graph. 1). Le fossile de Minatogawa 1 est
remarquable car il possède des foramens doubles de chaque côté. Les autres
Hommes pléistocènes ont des foramens doubles asymétriques, il s’agit de Mumba
10, Brno 2, Chancelade et Lagoa Santa parmi les Hommes anatomiquement modernes.
Le seul Néandertalien possédant un foramen mastoïdien double est Tabun 1. Aucun
foramen multiple n’apparaît chez les Pré-néandertaliens. Cf. Tab. 7
Symétrie
Les fréquences par côté sont plus faibles que les
fréquences individuelles. Cf. Tab. 2, 4 et 6. Ce déséquilibre est significatif
pour les foramens condylaires et pariétaux des Néandertaliens du Proche-Orient.
Les foramens asymétriques sont plus nombreux que les symétriques pour les
foramens mastoïdiens de l’échantillon du Gabon. Aucune différence n’apparaît
dans l’échantillon du Groenland. Des déséquilibres s’observent pour les
foramens condylaires, mastoïdiens, occipitaux et pariétaux des Hommes actuels.
Les différences entre les fréquences d’apparition
individuelles et par côté des échantillons d’hominidés qui vivent dans des
régions froides (Pré-néandertaliens, Néandertaliens d’Europe et Groenlandais)
ne sont jamais significatives. Au sein
d’échantillons de sujets de régions chaudes : Néandertaliens du
Proche-Orient et Gabonais, on remarque
pour certains foramens émissaires un déséquilibre significatif.
L’asymétrie est significative dans l’échantillon de
Néandertaliens du Proche-Orient pour les foramens condylaires et pariétaux,
alors que pareil déséquilibre est observé chez les sujets du Gabon uniquement
pour les foramens mastoïdiens. Il semble donc que selon la lignée d’hominidés,
le type de foramen le plus sensible à un déséquilibre varie. Cela indique t-il
des seuils de tolérance différents selon le type d’hominidé ?
Conclusion
Il se dégage certaines tendances entre les relations
du drainage émissaire, l’évolution humaine et l’environnement. Il faut les
nuancer notamment pour des raisons méthodologiques, comme sur la première
analyse en composantes multiples, par exemple, où la variance est assez faible
et où de nombreux sujets ont été exclus.
Il semble que l’augmentation du drainage émissaire
d’un point de vue quantitatif et qualitatif soit plus sensible au cours du
temps que selon un gradient thermique. Le nombre de foramens émissaires
augmente avec l’évolution, ainsi que celui du nombre de foramens multiples, à
l’exception des foramens rétro-articulaires. Les foramens symétriques sont plus
fréquents chez les Hommes actuels que chez les fossiles. Le nombre de foramens
uniques ou multiples, ainsi que l’asymétrie augmentent dans les milieux chauds.
Ce gradient est moins net que celui de l’évolution. Le froid semble favoriser
la stabilité du développement et ne nécessite pas l’apparition de nouvelles
veines émissaires qui protègent le cerveau de l’hyperthermie.
Remerciements: Je remercie
Madame Seurin ainsi que Messieurs Braga et Houët du laboratoire d’Anthropologie
des Populations du Passé de l’Université de Bordeaux 1, UMR 51989 PACEA, pour
leurs précieux conseils.
Bibliographie
Alemseged, Z., Coppens, Y., et
Geraads, D., 2002, Hominid cranium from Omo: description and taxonomy of
Omo-323-1976-896. Am. J. Phys. Anthropol., 117, 103-112.
Anton, S., 1997, Endocranial
hyperostosis in Sangiran 2, Gibraltar 1 and Shanidar 5, Am. J. Phys. Anthrop.,
102, 111-122.
Arsuaga, J.L., Martinez, I.,
Lorenzo, C., Gracia, A., Munoz, A., Alonso, O., et Gallego, J., 1999, The human
cranial remains from Gran Dolina lower pleistocene site (Sierra de Atapuerca,
Spain). J. Hum. Evol., 37, 431-457.
Arsuaga, J.L., Martinez, I., et
Gracia, A., 2001, Analyse phylogénétique des hominidés de la Sierra de
Atapuerca (Sima de los Huesos et Gran Dolina TD-6): l’évidence crânienne.
L’Anthropologie, 105, 161-178.
Bermudez de Castro, J.M., Arsuaga,
J.L., Carbonell, E., Rosas, A., Martinez, I., et Mosquera, M., 1997, A hominid
from the lower pleistocene of Atapuerca, Spain: possible ancestor to
Neandertals and modern humans. Science, 276, 1392-1395.
Bermudez de Castro, J.M.,
Martinon-Torres, M., Carbonell, E., Sarmiento, S., Rosas, A., van der Made, J.,
et Lozano, M., 2004, The Atapuerca sites and their contribution to the
knowledge of human evolution in Europe. Evolutionary Anthropology, 13, 25-41.
Bonfiglioli, B., Mariotti, V.,
Facchini, F., Belcastro, MG., et Condemi, S., 2004, Masticatory and
non-masticotary dental modifications in the epipaleolithic necropolis of
Taforalt (Morocco). Int. J. osteoarcheol., 14, 448-456.
Boyd, G.I., 1934, The emissary
foramina of the cranium in primates. Journal of Anatomy, 69 : 113-117
Braga, J., 1995. Définition de
certains caractères discrets crâniens chez Pongo, Gorilla et Pan, Perspectives
taxonomiques et phylogénétiques. Thèse de doctorat Université Bordeaux 1. pp. 398.
Braga, J., et Boesch, C., 1997a,
Further data about venous channels in South African Plio-Pléistocène hominids.
J. Hum. Evol. , 33, 423-447.
Braga, J., et Boesch, C., 1997b,
The «radiator» bias. A reply to Falk & Gage. J. Hum. Evol.,
33, 503-506.
Cabanac, M., et Brinnel, H., 1985,
Blood flow in the emissary veins of the human head during hyperthermia.
European Journal of Physiology, 54, 172-176.
Cabanac, M., 1986, Keeping a cool
head. News in Physiological Sciences, American Physiological Society, 1,
41-44
Coutselinis, A., Dritsas, C., et
Pitsios, Th., 1991, Expertise médico-légale du crâne pléistocène lA01/S2
(Apidima II), Apidima, Laconie, Grèce. L’Anthropologie, 95, n°2/3, 401-408.
Crubézy, E., Telmon, N., Sevin,
A., Picard, J., Rougé, D., Larrouy, G., Braga, J., Ludes, B., et Murail, P.,
1999, Microévolution d’une population historique. Étude des caractères discrets
de la population de Missimina (Soudan, III-VI siècle). Société d’Anthropologie
de Paris, 11, n°1-2, 7-200.
Dean, D., Hublin, J.J., Holloway,
R., et Ziegler, R., 1998, On the phylogenetic position of the pre-Neandertal
specimen from Reilingen, Germany. J. Hum. Evol., 34, 485- 508.
Falguères, C., Bahain, J.J.,
Yokoyama, Y., Arsuaga, J.L., Bermudez de Castro, J.M., Carbonell, E., Bischoff,
J.L., et Dolo, JM., 1999, Earliest humans in Europe: the age of TD 6 Gran
Dolina, Atapuerca, Spain. J. Hum. Evol., 37, 343-352.
Falk, D., 1986, Evolution of
cranial blood drainage in hominids: enlarged occipital/marginal sinuses and
emissary foramina. Am. J. Phys. Anthop., 70, 311- 324.
Falk, D., 1990, Brain evolution in
Homo: the «radiator» theory. Behavioral Brain Sciences, 13, 333-344.
Gray, H., 1973, Anatomy of the
human body, (29th American ed, Philadelphia, Lea and Febiger).
Grün, R., 1996, A re-analysis of
electron spin resonance dating results associated with the Petralona hominid.
J. Hum. Evol., 30, 227-241.
Hanihara, T., et Ishida, H., 2001,
Frequency variations of discrete cranial traits major human population. IV.
Vessel and nerve related variations. Journal of Anatomy, 199, 273-287.
Henry-Gambier, D., 2002, Les fossiles
de Cro-Magnon (les Eysies-de-Tayac, Dordogne) : nouvelles données sur leur
position chronologique et leur attribution culturelle. Paléo.,14, 201-204.
Holloway, R.L., 1990, Falk’s
radiator hypothesis. Behavioral Brain Sciences, 13, 360.
Iacumin, P., Cominotto, D., et
Longinelli, A., 1996, A stable study of mammal skeletal remains of
mid-pleistocene age, Arago cave, eastern Pyrennes, France. Evidence of
taphonomic and diagenetic effect. Paleogeography, Paleoclimatology,
Paleoecologie, 126, 151-160.
Kamminga, J., et Wright, R.V.S.,
1988, The Upeer cave at Zoukoudian and the origin of the Mongoloids. J. Hum.
Evol., 17, 739-767.
Leroyer, C., 1988, Des occupations
Castelperroniennes et Aurignaciennes dans leur cadre chrono-climatique. Dans La
mutation, édité par J.K. Kozlowski (Etudes et recherches archéologiques de
l’Université de Liège) p. 103-108.
Manzi, G., Vienna, A., et Hauser,
G., 1996, Developmental stress and cranial hypostosis by epigenetic trait
occurrence and distribution : an exploratory study on the italian neandertal,
J. Hum. Evol. 30, 511-527.
Minugh-Purvis, N., Radovcic, J.,
Smith, F.H., 2000, Krapina 1: a juvenile neandertal from the early late
pleistocene of Croatia. Am. J. Phys. Anthrop., 111, 393-424.
Oakley, K.P., 1964, The problem of
man’s antiquity. An historical survey. Bull. Br. Mus. Nat. Hist.Geology,
9
(5).
Pélissier-Hermitte, B., nd,
Approche méthodologique de quantification de certaines veines émissaires
cérébrales. Bull. et Mém. Soc. d’Anthrop. de Paris, (sous
presse).
Rink, W.J., Schwarcz, H.P., Lee,
H.K., Rees-Jones, J., Rabinovich, R., et Hover, E., 2001, Electron Spin
Resonance (ESR) and Thermal Ionization Mass Specrometric (TIMS) 230Th/ 234U
dating of teeth in middle paleolithic layers at Amud cave, Israel.
Geoarcheology, 6, 701-717.
Saban, R., 1984, Anatomie et
évolution des veines méningées chez les Hommes fossiles. E.N.S.B. Paris Mém.
Séc. Sci. 11, pp. 289.
Scheibel, A.B., 1990, Boiling over
in the great rift valley. Behavioral. Brain Sciences, 13, 364.
Schwarcz, H.P.,
Grün, R., Vandermeersch, B., Bar-Yosef, O., Valladas, H., et Tchernov, 1988,
ESR dates for the hominid site of Qafzeh in Israel. J. Hum. Evol., vol. 17,
n°8, 733-737.
Schwarcz, H.P., Simpson, J.J., et
Stringer, C.B., 1998, Neanderthal skeleton from Tabun: U-series data by
gamma-ray spectrometry. J. Hum. Evol., 35, 635-645.
Shea, J.J., 2003, Neandertals,
competition, and the origin of modern humans behavior in the levant.
Evolutionary Anthropology, 12, 173-187.
Silver, W.G., 1995, Cranial
emissary foramina and their relationship to climate. Am. J. Phys. Anthrop., 20
suppl., 195.
Trinkaus, E., Svoboda, J., West,
D.L., Sladek, V., Hillson, S.W., Drozdova, E., etFisakova, M., 2000, Human
remains from the moravian gravettian: morphology and taphonomy of isolated
elements from Dolni Vestonice II site. Journal of Archeological Science, 27, 1115-1132.
www. archaeologyinfo.com
www.modernhumanorigins.com
Zenker, W., et Kubik, S., 1996,
Brain cooling in human - anatomical consideration. Anat. Embryol., 193, 1-13.