
Colloques du Groupement des Anthropologistes de Langue Française (GALF)
Pouydebat, E., Gorce, P., Berge, C., Coppens, Y., 2006, La préhension chez les Primates: précision, outils et évolution. Antropo, 11, 159-166. www.didac.ehu.es/antropo
La préhension chez les Primates: précision, outils et évolution
Grasping among Primates: precision, tools and evolution
Emmanuelle Pouydebat1, Philippe Gorce2, Christine Berge1, Yves Coppens3
1 Laboratoire d’Anatomie comparée, M.N.H.N., USM 302, 55 rue Buffon, 75005 Paris
2 L.E.S.P. Université de Toulon et du Var, avenue de l’université, 83000 La Garde
3 Chaire de Paléoanthropologie, Collège de France, 75005 Paris
Correspondance: Emmanuelle Pouydebat. Laboratoire d’Anatomie comparée, M.N.H.N., USM 302, 55 rue Buffon, 75005 Paris. E-mail: epouy@wanadoo.fr
Mots clés: préhension, outil, précision, Primates, évolution
Keywords: grasping, tool, precision, Primates, evolution
Résumé
La préhension dite de "précision", impliquant les phalanges distales du pouce et de l’index, est parfois associée au genre humain (Napier, 1956), à l’usage d’outils (Susman, 1998) et à des critères morphologiques. Ces derniers, identifiés sur les fossiles, sont utilisés pour conclure à leur appartenance au genre humain et à leur capacité à employer des outils (Marzke, 1997). Sur la base d’observations de 69 spécimens d’humains, de Catarrhiniens et d’un Platyrrhinien, nous quantifions les zones des doigts utilisées au cours de tâches de saisies simples de petits et de gros objets, ainsi que de tâches complexes de proto-usage et d’utilisation d’outils. Nous concluons que la précision n’est pas le propre des humains et qu’elle n’est pas systématiquement liée à l’utilisation d’outils. Ceci nous permet de remettre en cause les critères morphologiques utilisés jusqu’à présent pour déduire la précision, l’utilisation d’outils et donc le caractère humain d’un fossile. De plus, nos analyses laissent apparaître une proximité récurrente entre les capucins et les humains, espèces éloignées d’un point de vue phylogénétique, suggérant l’existence possible de fonctions et de comportements proches malgré de grandes distinctions génétiques.
Abstract
The term "precision", defined as the ability to oppose the tips of thumb and index is generally attributed to humans, tool use and associated with morphological criteria. Identified in fossils, those criteria are used to assert that they belong to the genus Homo and they manipulated tools. Observations of 67 Catarrhines and one Platyrrhine allowed us to quantify surfaces of fingers used during simple tasks of grasping both small and large objects, as well as during complex tasks of proto-tool and tool use. We concluded that precision handling is not peculiar to humans and that it is not systematically linked to tool use. These results allowed us to discuss morphological traits used till now to deduce precision, tool use and hence the human character of a fossil. Besides, our analyses let appear a recurrent proximity between the capuchins and the humans, species distant in a phylogenetic point of view, suggesting the existence possible of functions and close behaviors in spite of great genetic distinctions.
Introduction
La préhension dite de « précision », impliquant les phalanges distales du pouce et de l’index, est souvent associée au genre humain (Napier 1956, 1960; Schultz 1969; Tuttle, 1965), à l’utilisation d’outils (Clark 1993; Marzke 1992; Susman 1979, 1998) et à des critères morphologiques. Ces derniers, identifiés sur les restes fossiles, sont utilisés pour conclure à leur appartenance au genre humain et à leur capacité à utiliser des outils (Marzke et al., 1992; Napier 1960).
La question principale posée par ce travail est de savoir si la précision est exclusivement humaine, si elle est liée à l’utilisation d’outils et si les espèces actuelles dépourvues des traits morphologiques correspondants sont réellement déficientes en terme de précision et d’utilisation d’outils, afin d’inférer les aptitudes préhensiles des fossiles. Dans ce but, nous développons une approche essentiellement basée sur des expérimentations et des observations, incluant une analyse des zones des doigts sollicitées en fonction d’une des propriétés fondamentales de l’objet (sa taille) et de la tâche accomplie.
Matériel
Les spécimens étudiés regroupent 69 primates humains et non humains observés en captivité, soit 9 capucins (Cebus apella), 14 chimpanzés (Pan troglodytes), 3 gorilles (Gorilla gorilla), 7 orangs outans (Pongo pygmaeus), 9 macaques (Macaca fuscata), 9 babouins (Papio papio), 9 enfants humains âgés de 2 à 5 ans (Homo sapiens) et 9 adultes humains (Homo s.). Les primates non humains observés ne possèdent pas les traits morphologiques habituellement associés à la précision et à l’utilisation d’outils (asymétrie des têtes de métacarpiens II et V, muscle long fléchisseur du pouce, muscles du pouce développés, pouce relativement long, large extrémité des phalanges distales, et pouce opposable en ce qui concerne les capucins).
Méthodes
Définition des zones des doigts
Les zones des doigts ont été définies suivant la terminologie anatomique classique : 1- pouce, 2- index, 3- majeur, 4- annulaire, 5- auriculaire; ex- extrémité; p- pulpe; d- distale; pr- proximale; lat- latérale; int- articulation interphalangienne; m- moyenne; med- médiale; paume- paume de main.
Protocole
Les zones des doigts impliquées dans la saisie de petits et de gros objets (leur dimension a été calibrée selon la taille des espèces) par les primates non humains ont été quantifiées selon la méthode de l’échantillonnage par sujet ("Focal animal sampling", Altman 1974). Chez les humains adultes et enfants, un protocole expérimental a été mis en place, au cours duquel chaque individu devait se saisir des objets posés sur une table. Au total, 7863 saisies ont été analysées.
Les zones des doigts impliquées dans la saisie de gros objets (noix ou noix de coco selon l’espèce) afin de les casser contre un support (proto-usage d’outils) ont été quantifiées selon la méthode de l’échantillonnage par sujet pour les capucins, suivant un enregistrement à volonté ("ad libitum sampling", Altman 1974) pour les gorilles et selon le protocole expérimental décrit ci-dessus pour les humains adultes. Au total, 1272 saisies ont été analysées.
Les zones des doigts impliquées dans la saisie de bâtons pour extraire de la nourriture (outils) ont été quantifiées selon un protocole expérimental comparable effectué pour les gorilles et les humains. Pour les gorilles, des bûches aménagées de trous étroits au fond desquels étaient écrasées des figues ont été disposées dans l’enclos avec des branches pourvues de feuilles. Les gorilles ont effeuillé ces branches pour s’en servir comme outils afin de récolter les fruits. Chez les humains, une petite bouteille en plastique, remplie de chocolat mou leur a été présentée avec une baguette en plastique dur. Les individus avaient pour unique consigne de goûter ce qui se trouvait dans la bouteille. En tout, 1298 saisies ont été analysées.
Statistiques
Les données ont fait l’objet d’une analyse de variance Anova de type I après avoir contrôlé les conditions d’application d’un test paramétrique (Lehner 1996). L’analyse de l’effet croisé de la taille et de l’espèce (facteurs fixes), puis de la tâche et de l’espèce (facteurs fixes), sur le pourcentage relatif de chaque zone de doigt (facteurs dépendants) a été effectuée. La valeur F (Fisher) a été calculée en divisant le carré moyen de chaque facteur (et de l’interaction) par le carré moyen résiduel ou erreur (Lehner 1996; Sokal et al., 1995). Une comparaison des moyennes deux à deux a ensuite été réalisée à l’aide d’un test de Newman-Keuls. Le seuil de signification usuel (p) a été sélectionné à 0,05. Nous avons également pris en compte le seuil de signification calculé d’après la correction de Bonferroni afin de limiter le risque d’erreur d’évaluation des résultats expérimentaux ("overall experiment wise error", Sokal et al., 1995).
Résultats
Nous présentons ici une partie des résultats. Dans un premier temps, nous avons pu constater que la préhension de petits objets engendrait en moyenne une plus forte variabilité de types de saisies (plus de 40 recensés par individu) que la préhension de gros objets (20 en moyenne), d’objets dans le cadre du proto-usage d’outils (1 type de saisie) et d’outils comme les bâtons (7 types en moyenne). Quelle que soit la taille de l’objet ou la tâche, les humains adultes se sont avérés être les plus "spécialisés" en terme de faible variabilité des modes de préhension utilisés.
Dans un deuxième temps, nous avons mis en évidence que toutes les espèces étudiées ont fait preuve de "précision" en utilisant les phalanges distales de leur pouce et de leur index pour saisir des petits objets, dans des proportions diverses (de 5% des types de saisie chez les chimpanzés à 52% pour les capucins contre 42% chez les humains adultes). Il a été particulièrement intéressant de mettre en valeur la proximité entre les capucins et les humains, malgré leur éloignement phylogénétique (Figure 1).
Dans un troisième temps, nous avons pu montrer que la taille de l’objet avait une incidence très significative sur les zones des doigts utilisées (Pouydebat et al., 2004), de même que la tâche. A cet égard, les humains adultes et les capucins utilisent leurs phalanges distales de manière quasi exclusive pour saisir les petits comme les gros objets. En outre, les extrémités du pouce et de l’index ne sont pas systématiquement impliquées dans les tâches complexes de proto-usage et d’utilisation d’outils. Afin de rendre compte de ces résultats, nous avons comparé les zones des doigts utilisées dans :
- la saisie de gros objets et de "proto-outils" par les humains adultes, les gorilles et les capucins (Figure 2)
- la saisie de bâtons pour pêcher par les humains et les gorilles (Figure 3).

Figure 1. Saisies de précisions utilisées par les humains et les capucins pour saisir de petits objets
Figure 1. Precision grasping used by humans and capuchins to grasp small objects

Figure 2. Saisies de gros objets et proto-usage d’outils. a: gorilles. b: capucins
Figure 2. Grasping of large objects and proto tool use. a: gorillas. b: capuchins
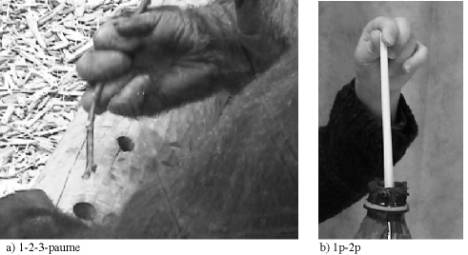
Figure 3. Exemples de zones de doigts utilisées pour pêcher. a: gorille. b: enfant
Figure 3. Examples of grasps used to extract food: a: gorilla. b: child
Les résultats montrent (Tableau 1) que la tâche a un effet très significatif sur les zones des doigts utilisées, quelle que soit l’espèce. En revanche, il ne semble pas exister de différence significative entre les espèces. Le test de Newman Keuls apporte quelques précisions. Ainsi, Gorilla g. n’utilise jamais les phalanges distales de son pouce, de son index et de son majeur.
|
Variables
|
Tâche F Dl = 1,42 |
Espèce F dl = 2,42 |
Tâche x Espèce F dl = 2,42 |
|
1p-2p-3p 1p-2p-3p-4p 1p-2p-3p-4p-5p 1-2-3-4-5-paume 1-2-3-paume |
58,911** 75,279** 102,650** 637,19** 17,748** |
1,482 1,249 1,001 0,198 3,957* |
143,432** 263,531** 1498,010** 3372,189** 591,785** |
Tableau 1. Effet de la tâche sur les zones de doigts, toute espèces confondues. Variables: zones des doigts (p: pulpe ; 1-5: doigt) ; Tâche: saisie simple de gros objets et saisie complexe de proto outils ; Espèces: 68 individus appartenant à 7 genres de primates humains et non humains ; F: test de Fisher ; Dl: degré de liberté ; * : seuil de significativité p < 0,05 ; ** : seuil de significativité p < 0,01 selon les corrections du test de Bonferroni)
Table 1. Task influence on the surface of the fingers for all species. Variables : surfaces of digits (p: pulp; 1-5: digit); Task (tâche): simple grasping of small and large objects and complex grasping of proto-tool use; Species (espèces): 68 specimens belonging to 7 genus of primates and non human primates; F: Fisher test; SD (dl): standard deviation; *: significant difference p < 0,05; **: significant difference p < 0,01 according to the corrections of the test of Bonferroni
Enfin, les zones des doigts utilisées par les humains adultes, enfants et les gorilles dans la saisie de bâtons pour extraire de la nourriture sont comparées. Les résultats de l’étude statistique montrent que l’espèce a un effet très significatif sur les saisies utilisées pour "pêcher". Ainsi, les enfants humains, les adultes humains et les gorilles ne saisissent pas les bâtons de la même manière. Toutes les zones des doigts présentent des différences significatives selon l’espèce (Figure 4).
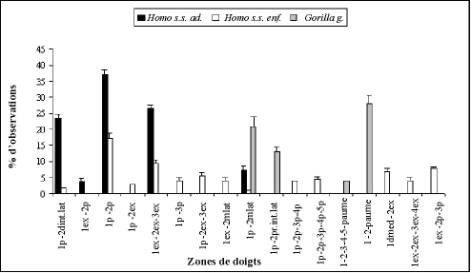
Figure 4. Zones de doigts utilisées par les humains adultes, enfants
et les gorilles pour saisir "pêcher" ; cette figure montre la polyvalence des enfants et la précision des adultes humains contrairement aux gorilles qui préconisent l’utilisation de leur paume et le bord latéral de leur index.
Figure 4. Surfaces of the fingers used by human adults, children and gorillas
to grasp a branch in order to extract food; this figure show the variability of grip types among children, the precision of human adults and the use of palm and lateral surfaces of the index by gorillas.
Les zones des doigts utilisées le plus fréquemment pas les humains concernent les pulpes distales du pouce et de l’index ainsi que l’extrémité des trois premiers doigts. Ce n’est pas le cas chez les gorilles qui pour cette même tâche accomplissent des saisies impliquant la paume ou le pouce et le bord latéral de l’index. Cette dernière saisie est également employée par les humains adultes qui, une nouvelle fois, sont susceptibles d’utiliser le bord latéral de leur index pour une tâche complexe.
Discussion
L’utilisation d’outils est fréquemment corrélée à la précision, dans la mesure où toutes deux nécessitent un effort cognitif et manuel considérable (Costello, Fragaszy 1988; Marzke et Wullstein, 1996). Ainsi, pour certains auteurs (Napier, 1960; Jones-Engels et Bard, 1996), la saisie de précision a probablement eu des implications dans l’évolution de l’organisation cérébrale et de l’intelligence. Si un fossile présente des dispositions morphologiques propres à la saisie de précision, alors il s’agit probablement d’un Hominidé appartenant à la lignée humaine qui utilisait des outils (Marzke 1997; Susman 1989, 1998). Notre travail montre cependant que la réalité est plus complexe. Tout d’abord, les primates non humains testés (Pan t., Pongo p., Gorilla g., Macaca f., Papio p. et Cebus a.) ont tous utilisé l’extrémité du pouce et de l’index, suggérant ainsi que les critères morphologiques (absents chez ces non humains) utilisés jusqu’ici pour déduire la précision sont à remettre en cause. D’autant plus que l’espèce la plus proche des humains s’est avérée être la capucin, dont le pouce est considéré comme pseudo-opposable. Cette proximité inattendue entre ces deux espèces éloignées d’un point de vue phylogénétique montre à ce titre que deux espèces peuvent partager des points communs fonctionnels et cognitifs malgré leurs différences génétiques. Il n’en reste pas moins que les humains se distinguent nettement des autres primates par une plus faible variabilité des types de saisies.
Par ailleurs, les saisies de "puissance" impliquant la paume ne doivent plus être nécessairement associées à un aspect primitif. En effet, ces saisies sont utilisées par les gorilles lorsque ceux-ci emploient des outils suivant une optimisation spatiale et temporelle caractéristique d’une activité cognitive élevée (Pouydebat et al., 2005). A l’inverse, la précision n’est pas systématiquement associée à l’outil. En effet, lorsque les gorilles effeuillent les branches et s’en saisissent pour "pêcher", ils privilégient des saisies puissantes, contrairement aux humains adultes qui adoptent des saisies précises. Autrement dit, les gorilles, très précis pour une tâche simple (saisie de petits objets), sont puissants dans une tâche complexe. De même, dans la manipulation de proto-outils et d’outils pour casser des noix, les humains (comme les gorilles et les capucins) utilisent les saisies puissantes. En outre, le chimpanzé, probablement l’espèce actuelle non humaine la plus créative et utilisatrice d’outils (Boesch et Boesch, 1990) est aussi celle (avec l’orang-outan) qui utilise le moins les saisies de précision (5 à 7% des saisies de petits objets). Cet aspect montre bien que l’on peut posséder des capacités manuelles développées (utilisation d’outils variés) sans pour autant être précis. A l’inverse, les babouins ou les macaques sont très précis mais présentent, jusqu’à preuve du contraire, des aptitudes à fabriquer des outils beaucoup moins développées que le chimpanzé. Enfin, deux saisies impliquant le bord latéral de l’index (1p-2mlat et 1ex–2dint.lat), sont utilisées dans une tâche complexe (pêche) par les humains. Ces saisies ne peuvent donc plus être considérées comme "inefficaces" (Napier 1960) ou "imprécises" (Costello et Fragaszy, 1988).
En terme d’interprétation, il est désormais insuffisant de déduire qu’un fossile est susceptible d’avoir utilisé des outils et donc qu’il est apparenté aux humains au seul regard du fait que sa main pouvait être précise. A l’inverse, une main qui ne semble pas avoir été précise n’est pas nécessairement dépourvue de capacité à fabriquer des outils. De plus, les critères morphologiques utilisés jusqu’à présent ne semblent pas pertinents pour démontrer la précision et l’utilisation d’outils. Par exemple, l’aptitude des capucins à être précis et à utiliser des outils fait de l’opposabilité du pouce un critère morphologique insuffisant. D’autant plus que les capucins ont su mettre en place une véritable stratégie d’utilisation des supports les plus durs afin de rendre plus efficace le cassage des noix (Pouydebat et al., 2005). La planification de l’action témoigne d’une complexité cognitive que nous ne serons malheureusement jamais en mesure d’interpréter via l’analyse morphologique des fossiles.
Conclusion
Ce travail montre, dans un premier temps, la nécessité d’intégrer la variabilité de la taille de l’objet et de la tâche dans l’étude de la préhension, dans la mesure où ces paramètres influencent les modes de saisie. Il montre également l’importance de s’extraire quelque peu de la dichotomie entre précision et puissance définie par Napier (Napier, 1956), si l’on prend en compte la variabilité des zones des doigts impliquées.
Par ailleurs, toutes les espèces testées se sont avérées précises, bien que certaines, comme les chimpanzés, le sont dans des proportions moindres que les autres. Ainsi, une espèce peut être peu précise (utiliser davantage le bord latérale de son index) et être fabricante et utilisatrice d’outils. De même, une tâche complexe peut être associée à la puissance (utilisation de la paume). A l’inverse, il ne suffit pas d’être précis pour être un grand fabricant d’outils. Ceci remet en question les études attestant que certains fossiles seraient dans la lignée humaine parce qu’ils ont les caractéristiques de la précision (corrélées à l’outil) alors que d’autres fossiles en sont écartés parce qu’il n’ont pas ces caractéristiques. Par ailleurs la proximité entre les capucins et les humains, récurrente dans nos travaux, montre bien les affinités fonctionnelles et cognitives susceptibles d’exister entre deux espèces éloignées d’un point de vue phylogénétique. Enfin, la complexité de la préhension ne peut se résumer aux surfaces des doigts mises en jeu. Nous pouvons cependant chercher les critères morphologiques réellement liés aux zones de doigts et développer d’autres méthodes inspirées de l’observation du vivant et de la biomécanique humaine, afin de mieux cerner les paramètres impliqués dans l’évolution des capacités manuelles des primates.
Remerciements. Les auteurs tiennent à remercier Monsieur José Braga pour son invitation au colloque du Galf (Toulouse 2005) ainsi que tout le personnel du zoo de Beauval et du parc de la vallée des singes.
Bibliographie
Altmann, G., 1974, Observational study of behavior. Sampling methods, Behavior, 49, 227-267.
Boesch, C. et Boesch, E., 1990, Tool use and tool making in wild chimpanzees. Folia Primatologica, 54, 86-99.
Clark, J.D., 1993, Stone artifact assemblages from Members 1-3 Swartkrans Cave. Dans Swartkrans: a cave’s chronicle of early man, édité par C.K. Brain (Transvaal Museum Memorial) p.167-194.
Costello, M.B. et Fragaszy, D.M., 1998, Prehension in Cebus and Saimiri : grip type and hand preference. American Journal of Primatology, 15, 235-245.
Jones-Engels, L. et Bard K.A., 1996, Precision grips in young chimpanzees. American Journal of Primatology, 39 (2), 1-15.
Lehner, P.N., 1996, Handbook of ethological methods, Cambridge University Press, p. 672.
Marzke, M.W., 1997, Precision grips, hand morphology and tools. American Journal of Physical Anthropology, 102, 91-110.
Marzke, M.W., Wullstein, K.L. et Viegas, S.F., 1992, Evolution of the power grip and its morphological correlates in hominids. American Journal of Physical Anthropology, 89, 283-298.
Marzke, M.W. et Wullstein, K.W., 1996, Chimpanzee and human grips: a new classification with a focus on evolutionary morphology. International Journal of Primatology, 17, 117-139.
Napier, J.R., 1956, The prehensile movements of the human hand. Journal of Bone and Joint Surgery, 38B, 902-913.
Napier, J.R., 1960, Studies of the hands of living primates. Proceeding of Zoological Society of London, 134, 647-657.
Pouydebat, E., Berge, C. et Gorce P., 2005, Fittings and use of branches as tools to extract food by captive gorillas. Folia Primatologica, 76, 180-183.
Pouydebat, E., Gorce, P. et Berge C., 2005, Substrate optimisation in nuts cracking by capuchin monkeys. American Journal of Primatology (In Press).
Pouydebat, E., Gorce, P., Berge, C. et Coppens Y., 2004, Biomechanical study of grip types among primates: object size influence. Archives of Physiology and Biochemistry, 112, p.117.
Schultz, A.H., 1969, The life of Primates, édité par Weidenfeld and Nicolson (London), p.341.
Sokal, R.R. et Rohlf, F.J., 1995, Biometry : the principles and practice of statistics in biological research, édité par Freeman and Company, p. 887.
Susman, R.L., 1979, Comparative and functional morphology of hominoid fingers. American Journal of Physical Anthropology, 50, 215-236.
Susman, R.L., 1989, New hominid fossils from the Swartkrans formation: postcranial specimens. American Journal of Physical Anthropology, 79, 451-474.
Susman, R.L., 1998, Hand function and tool behavior in early hominids. Journal of Human Evolution, 35, 23-46.
Tuttle, R.H., 1965, A study of the chimpanzee hand with comments on hominoid evolution, University of California, Berkeley, p. 526.