
Salces, I., Rebato, E., Rosique, J., Slachmuylder, J. L., Vercauteren. M. y Susanne, C., 2001. Fuentes genéticas y ambientales en la transmisión familiar del índice de Quetelet. Antropo, 0, 61-69 www.didac.ehu.es/antropo
Fuentes genéticas y ambientales en la transmisión familiar del índice de Quetelet
Itziar Salces1, Esther Rebato1, Javier Rosique1, Jean-Louis Slachmuylder2, Martine Vercauteren2, Charles Susanne3
1Universidad del País Vasco, 2Université Libre de Bruxelles, 3Vrije Universiteit Brussels
Dirección para correspondencia: Itziar Salces. Laboratorio de Antropología Física. Dpto. Biología Animal y Genética. Facultad de Ciencias. Universidad del País Vasco. Apdo. 644. 48080-Bilbao. España. E-mail: ggbsabei@lg.ehu.es
Palabras clave : Indice de Quetelet, Transmisión familiar, Análisis path, Familias, País Vasco.
Se estudió una muestra transversal de 1024 individuos (115 padres, 257 madres, 309 hijos y 343 hijas) pertenecientes a 442 familias nucleares residentes en la provincia de Bizkaia (País Vasco). El objetivo fue estimar los componentes transmisibles (genéticos y culturales) y no transmisibles que están actuando sobre la semejanza familiar del índice de Quetelet, por medio de un análisis path tipo BETA. Se probaron diversos modelos de transmisión familiar para averiguar cuál ajustaba mejor la transmisión de padres a hijos, a fin de estimar la importancia relativa que los factores genéticos, los culturales transmitidos y los ambientales no transmisibles, tienen sobre la herencia de esta variable. Los resultados han mostrado que el modelo general, estimador de 10 parámetros, es el que mejor ajusta la transmisión del índice de Quetelet. Se han encontrado, asimismo, evidencias de una posible influencia materna sobre este índice, así como estimas del componente transmisible total más bajas que las publicadas en la literatura, si bien hay que tener presente que estas estimas son específicas para cada población.
A cross-sectional sample of 1024 individuals (115 fathers, 257 mothers, 309 sons and 343 daughters) of 442 nuclear families living in the province of Biscay (Basque Country) was studied. The aim was the estimation of the transmissible (genetic and cultural) and non-transmissible components that are acting on the familial resemblance for the Quetelet9s index, through a BETA model of path analysis. In addition, several models of familial transmissible were tested, in order to find out what was that best fitting the transmission from parents to their children, and to estimate the relative importance of the genetic, cultural transmitted and environmental non-transmitted factors have on the inheritance of this variable. The results have showed that the full model, which estimated 10 parameters, was the lest restrictive model that best fitted the Quetelet9s index transmission. Also, evidences of a possible maternal influence on this index, as well as estimations of the total transmissible component lower than those in the literature were found. However, we could not forget that these estimations are specific for each population.
La mayoría de los estudios realizados sobre la herencia de los caracteres de variación continua se ha basado en el análisis de correlaciones entre individuos biológicamente emparentados. Los rasgos más frecuentemente estudiados han sido la estatura y el peso (Rao et al. 1975, Sánchez-Andrés y Mesa 1994, Rebato et al. 1997, Salces et al. 2001a,b). Otros temas de interés han sido la genética de la obesidad, considerada desde puntos de vista muy diferentes (pe. Sørensen et al. 1989), o las influencias genéticas y ambientales sobre diversas medidas corporales (Mueller y Malina 1980, Roberts 1985), entre otros. Todas estas investigaciones han intentado aproximarse a una mejor comprensión de la variación fenotípica sobre la transmisión de estos caracteres entre padres e hijos, y a cómo la contribución genética puede alterarse por cambios en las condiciones ambientales y el estilo de vida de los individuos y las poblaciones. Durante las 3 últimas décadas, algunos investigadores han utilizado la técnica del análisis path (Wright 1921) para analizar no solamente la semejanza entre los individuos, sino el papel jugado por los factores genéticos y ambientales. Estos factores, que se transmiten entre generaciones, están actuando junto con los factores ambientales no transmisibles sobre los fenotipos de los individuos y sobre la semejanza familiar entre los emparentados (Rao et al. 1982, 1984, Rice et al. 1978, Cloninger et al. 1979a,b).
La presente investigación se emprendió con el objetivo de estimar los componentes transmisibles (genéticos y culturales) y no transmisibles que pueden estar actuando sobre la semejanza familiar del índice de Quetelet, en una muestra poblacional de familias nucleares residentes en la provincia de Bizkaia (País Vasco). Asimismo, se probaron diferentes modelos de transmisión familiar con el objeto de buscar cuál era el que mejor ajustaba la transmisión de padres a hijos en la muestra estudiada, y estimar la importancia relativa que los factores genéticos, los culturales transmisibles y los ambientales no transmitidos, tienen sobre la herencia de este índice ponderal.
Se estudió, por metodología transversal, una muestra familiar compuesta por 1024 individuos (115 padres, 257 madres, 309 hijos y 343 hijas) pertenecientes a 442 familias nucleares residentes en la provincia de Bizkaia (País Vasco). El rango de edad fue de 22 a 66 años entre los padres, de 22 a 58 años en las madres, de 4 a 22 los chicos y de 4 a 27 años las chicas. La edad decimal de cada individuo se calculó como la diferencia entre el día del muestreo y la fecha de nacimiento. Se midieron la estatura (cm) y el peso (kg) siguiendo los criterios del Programa Biológico Internacional (Weiner y Lourie 1981), y se calculó el índice de Quetelet (peso (kg)/estatura (m2)) o índice de masa corporal (IMC), que es un índice de robustez de gran relevancia sanitaria y epidemiológica, pues se ha comprobado que está correlacionado con la composición corporal (Mueller 1983, Longini et al. 1984), además de ser un interesante indicador del sobrepeso y la obesidad (OMS 1995). Los sujetos respondieron a un cuestionario en el que, entre otros datos, se les preguntaba qué alimentos (sólidos y/o líquidos) componían su desayuno habitual, si es que acostumbraban tomar alguno. Sólo se consideró como respuesta del cuestionario Cno desayunaD la de aquellas personas que regularmente tenían por hábito no ingerir nada en las primeras horas de la mañana o incluso hasta el mediodía.
Para
realizar las comparaciones entre los diferentes tipos de parientes, evitar los posibles
efectos de la evolución secular y la influencia de la involución senil
(Susanne 1974, 1977) sobre los rasgos estudiados, se procedió a estandarizar
los datos de forma independiente en las dos generaciones (parental y descendientes)
para cada sexo. La submuestra parental se dividió, además de en función
del sexo, en dos grupos de edad: <46 años y e46 años. A continuación,
se calcularon las puntuaciones estándar o SDS para cada individuo, utilizando
los valores de la media y la desviación estándar para cada subgrupo, según
la fórmula ![]() . En cuanto
a los valores medios de referencia en las submuestras de descendientes, éstos
se estimaron a través de la ecuación de regresión que mejor ajustaba
la nube de puntos, por grupos de edad y sexo separadamente. Por su parte, las desviaciones
estándar de referencia se obtuvieron a partir del ajuste de las desviaciones
estándar calculadas para grupos de edad de un año de los descendientes.
Estas curvas (en ambos casos polinomios de 2º grado) proporcionaban estimaciones
adecuadas de la media y desviación estándar muestrales a edades específicas
de los sujetos estudiados. Los SDS se utilizaron para calcular todos los coeficientes
de correlación así como para el análisis path.
. En cuanto
a los valores medios de referencia en las submuestras de descendientes, éstos
se estimaron a través de la ecuación de regresión que mejor ajustaba
la nube de puntos, por grupos de edad y sexo separadamente. Por su parte, las desviaciones
estándar de referencia se obtuvieron a partir del ajuste de las desviaciones
estándar calculadas para grupos de edad de un año de los descendientes.
Estas curvas (en ambos casos polinomios de 2º grado) proporcionaban estimaciones
adecuadas de la media y desviación estándar muestrales a edades específicas
de los sujetos estudiados. Los SDS se utilizaron para calcular todos los coeficientes
de correlación así como para el análisis path.
Para construir el índice ambiental que posteriormente se introducirá en el análisis path se empleó un análisis de correspondencias múltiples, capaz de analizar variables categóricas u ordinales. Este análisis trata exclusivamente variables nominales, asignando valores numéricos a casos (o sujetos) y categorías, denominados puntuaciones de los objetos y cualificaciones, respectivamente. Los datos sobre los que se aplicó este análisis fueron las respuestas obtenidas en la encuesta realizada a los sujetos estudiados, referente a qué alimentos componían su desayuno habitual (Tabla 1). Las puntuaciones obtenidas de los objetos fueron los datos individuales que representan el consumo de alimentos en el desayuno, y que constituyen el índice ambiental para el análisis path.
|
Variables/Categorías |
1 |
2 |
3 |
4 |
|
Mermelada-miel |
944 |
80 |
|
|
|
Bollería |
863 |
161 |
|
|
|
Galletas |
682 |
342 |
|
|
|
Leche |
73 |
951 |
|
|
|
Cereales |
878 |
146 |
|
|
|
Huevos-embutidos |
1010 |
14 |
|
|
|
Pan |
855 |
169 |
|
|
|
Zumos-Fruta |
861 |
163 |
|
|
|
Bebidas calientes (Achicoria-Café-Cacao) |
386 |
6 |
290 |
342 |
Tabla 1. Variables categóricas del desayuno. 1: No consume, 2: consume, salvo en Bebidas Calientes donde 1: No consume, 2: Achicoria, 3: Café, 4: Cacao soluble.
Table 1. Categorical variables of breakfast consumption. 1. Do not take, 2. Do take, except for Hot drinks where 1. Do not take, 2. Chicori, 3. Coffee, 4. Cocoa.
Como método de análisis estadístico se ha empleado el denominado PATHMIX, que es un caso especial de análisis path tipo BETA (Figura 1). Dicho método permite estimar correlaciones dentro de familias nucleares y/o ajustar modelos de análisis path a datos familiares, considerando las estimas de varias correlaciones familias como independientes, aunque sean de los mismos datos (Cloninger et al. 1979a,b). En esta investigación las correlaciones se han estimado individualmente, en contraposición con la estimación simultánea realizada por Rao et al. (1979), lo que evita encontrar los problemas numéricos asociados a las estimas simultáneas (Rao et al. 1982). Además, PATHMIX tiene la ventaja de que impide que las familias con un mayor número de hijos introduzcan más información. Para ello estima un tamaño muestral teniendo en cuenta la información independiente aportada por cada nuevo hermano de la fratria. Los tamaños muestrales estimados se calculan invirtiendo la varianza asintótica de las correlaciones calculadas (Rao et al. 1982), que son las que aparecen en la Tabla 2.
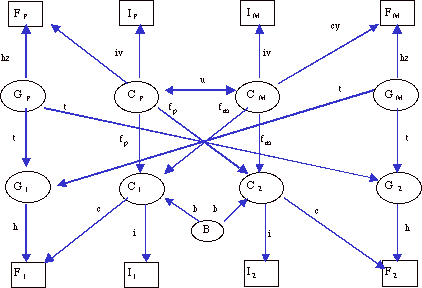
Figura 1. Modelo BETA de anális path utilizado para el análisis de la muestra familiar vizcaína (G = Genotipo, F = Fenotipo, I = Indice, C = Ambiente transmisible, B = Factores socio-culturales no transmisibles, P = padre, M = Madre, 1 y 2 = Descendientes).
Figure 1. BETA model of path analysis used for the analysis of the Biscaian familial sample (G = Genotype, F = Phenotype, I = Index, C = Transmissible environment, B = Non- transmissible socio-cultural factors, P = Father, M = Mother, 1 and 2 = Offspring).
Tras
calcular las correlaciones esperadas (Tabla 2), se puede determinar el modelo de
transmisión familiar que se va a aplicar a los datos (la hipótesis contrastada).
Se contrastó el modelo general, con el que se estimaban los l0 parámetros
que se muestran en la Tabla 3, frente a 4 modelos específicos, que se detallan
en la Tabla 4. En dicha tabla aparece, igualmente, el número de parámetros
fijados en cada uno de los modelos de transmisión y los grados de libertad del
test de ![]() .
.
|
|
Correlación esperada |
|
Correlación esperada |
|
Fenotipos |
|
Indices |
|
|
FPFM |
|
IPIM |
|
|
FPFH |
|
IPIH |
|
|
FMFH |
|
IMIH |
|
|
FH1FH2 |
|
IH1IH2 |
|
|
Entre rasgos |
|
Entre rasgos |
|
|
FPIM = IPFM |
ucyiv |
IMFH |
|
|
FPIH |
|
FPIP = FMIM |
cuiv |
|
IPFH |
|
FH1IH2 = FH2IH1 |
|
|
FMIH |
|
FHIH |
ci |
Tabla 2. Correlaciones esperadas que pueden estimarse con el método PATHMIX de análisis path (F = Fenotipo, I = Indice, P = padre, M = Madre, H1 y H2 = Descendientes).
Table 2. Expected correlations which could be estimated through the PATHMIX method of path analysis (F = Phenotype, I = Index, P = Father, M = Mother, H1 and H2 = Offspring).
|
Parámetro |
Definición |
|
h |
Efecto del genotipo infantil sobre el fenotipo infantil |
|
z |
(h . z) = efecto del genotipo adulto sobre el fenotipo adulto |
|
c |
Efecto del ambiente transmisible infantil sobre el fenotipo infantil |
|
y |
(c . y) = efecto del ambiente transmisible adulto sobre el fenotipo adulto |
|
u |
Correlación entre los ambientes transmisibles parentales adultos a través de homogamia social |
|
ff |
Efecto del ambiente transmisible del padre sobre el del hijo que cría, ambos ambientes medidos simultáneamente |
|
fm |
Efecto del ambiente transmisible de la madre sobre el del hijo que cría, ambos ambientes medidos simultáneamente |
|
b |
Efecto del ambiente común no transmisible de la fratria sobre el ambiente del hijo |
|
i |
Efecto del ambiente transmisible infantil sobre el índice del hijo |
|
v |
(i . v) = efecto del ambiente transmisible progenitor sobre su índice |
Tabla 3. Parámetros estimados con el método PATHMIX de análisis path.
Table 3. Parameters estimated with the PATHMIX method of path analysis.
Los fenotipos estandarizados del índice de Quetelet, así como el índice ambiental construido, generaron 16 correlaciones. Dichas correlaciones, junto con los tamaños muestrales correspondientes y sus errores estándar, se presentan en la Tabla 5.
|
Modelo de transmisión |
Hipótesis |
Nº parám. fijados |
g.l. |
|
General |
|
0 |
6 |
|
Sin homogamia social |
u = 0 |
5 |
1 |
|
Sin transmisión cultural |
b = 0 |
5 |
1 |
|
Sin efectos genéticos |
h = 0 |
3 |
2 |
|
Más reducido |
c = y = 0; i = v = 0 |
2 |
4 |
Tabla 4. Cuadro resumen de los modelos de transmisión familiar contrastados, así como el número de parámetros fijados en cada uno y los correspondientes grados de libertad en el test del cociente de la probabilidad.
Table 4. Summary of the tested models of familial transmission, as well as the number of fixed parameters in each one and the corresponding degrees of freedom in the likelihood ratio test.
|
|
FP-FM |
FP-FH |
FM-FH |
FH1-FH2 |
IP-IM |
IP-IH |
IM-IH |
IH1-IH2 |
|
n |
92 |
105 |
239 |
75 |
92 |
105 |
239 |
75 |
|
r |
0,052 |
0,120 |
0,240*** |
0,225* |
0,295** |
0,156* |
0,094 |
0,127 |
|
es |
0,121 |
0,080 |
0,056 |
0,105 |
0,086 |
0,077 |
0,058 |
0,140 |
|
|
|
|
|
|
|
|
|
|
|
|
FP-IM |
FP-IH |
IP-FH |
FM-IH |
IM-FH |
FP-IP |
FH1-IH2 |
FH-IH |
|
n |
92 |
105 |
105 |
239 |
239 |
106 |
253 |
253 |
|
r |
-0,034 |
0,042 |
-0,072 |
0,023 |
-0,065 |
-0,105 |
0,014 |
-0,006 |
|
es |
0,076 |
0,075 |
0,082 |
0,060 |
0,059 |
0,053 |
0,087 |
0,055 |
Tabla 5. Estimas, por máxima verosimilitud, de las correlaciones familiares (r) para el índice de Quetelet, sus tamaños muestrales (n) y los errores estándar de las correlaciones (es). Se indica, asimismo, la significación cuando existe. (*: p£ 0,05; **: p£ 0,01; ***: p£ 0,001; F: Fenotipo, I: Indice, P: Padre, M: Madre, H: Hijos).
Table 5. Familial correlations (r) estimated through maximum likelihood for the Quetelet9s Index, sample sizes (n) and standard errors of the correlations (es). The signification, when it existed, was also showed. (*: p£ 0,05; **: p£ 0,01; ***: p£ 0,001; F: Phenotype, I: Index, P: Father, M: Mother, H: Children).
Los parámetros estimados para la transmisión familiar de las variables estudiadas, bajo todos los modelos contrastados, así como los resultados del test de bondad de ajuste de dichos modelos de transmisión, aparecen en la Tabla 6. En dicha tabla se puede observar cómo el modelo general de transmisión se ha ajustado considerablemente bien (p d 0,001) para la transmisión familiar del índice de Quetelet en la muestra familiar vasca. El parámetro h (raíz cuadrada de la heredabilidad en sentido amplio, h2) presentó un valor de 0,53 bajo el modelo general de transmisión familiar.
|
TEST DE HIPOTESIS |
|
|
PARAMETROS |
|||||||||||
|
Modelo |
g.l. |
c2 |
p |
|
h |
c |
y |
z |
u |
ff |
fm |
b |
i |
v |
|
General |
6 |
26,30 |
*** |
|
0,533 |
0,000 |
0,855 |
1,006 |
0,510 |
0,215 |
0,221 |
0,505 |
0,086 |
0,878 |
|
u = 0 |
1 |
0,35 |
|
|
0,536 |
0,000 |
0,858 |
1,006 |
0,028 |
0,208 |
0,226 |
0,505 |
0,027 |
0,856 |
|
b = 0 |
1 |
0,07 |
|
|
0,535 |
0,000 |
0,855 |
1,005 |
0,510 |
0,215 |
0,223 |
0,000 |
0,087 |
0,878 |
|
h = z = 0 |
2 |
2,48 |
|
|
0,001 |
0,473 |
0,973 |
0,000 |
0,613 |
0,404 |
0,456 |
0,557 |
0,000 |
0,925 |
|
c = y = 0, i = v = 0 |
4 |
6,75 |
|
|
0,500 |
0,000 |
0,000 |
1,000 |
0,499 |
0,199 |
0,198 |
0,500 |
0,997 |
0,998 |
Tabla 6. Modelos contrastados, tests de hipótesis y parámetros estimados bajo cada modelo para el Indice de Quetelet (***: p£ 0,001).
Table 6. Tested models, tests of the hypothesis and estimated parameters under each model for the Quetelet9s Index (***: p£ 0,001).
Las estimas de los parámetros fP y fM (ambientes transmisibles paterno y materno, respectivamente, sobre sus hijos) pueden reflejar posibles efectos diferenciales de los progenitores, actuando sobre la transmisión familiar de la variable estudiada. Así, en la Tabla 6 se puede observar como fM es, en términos generales, superior a fP bajo todos los modelos de transmisión planteados. Ninguno de los 4 submodelos de transmisión (Tabla 6) contrastados con el modelo general se han ajustado para la transmisión familiar del índice de Quetelet.
El estudio de los componentes de la varianza (Tabla 7) muestra una contribución de los factores culturales transmisibles (b2) del 22% para el índice de Quetelet bajo el modelo de transmisión sin efectos genéticos. Bajo el resto de modelos, la transmisibilidad total (t2) de la varianza fenotípica observada se sitúa en torno al 28%. El modelo de transmisión mas reducido (c = y = 0, i = v = 1) ha puesto de manifiesto una fracción transmisible de la varianza (t2) del 25%, mientras que el 75% restante sería debido a la varianza ambiental no transmisible, pero que podría ser compartida por los individuos de la misma generación (hermanos).
|
|
Varianza transmisible |
|
Varianza |
||
|
|
Total |
Biológica |
Cultural |
ambiental |
|
|
Modelo |
t2 |
h2 |
b2 |
|
e2 |
|
General |
0,284 |
0,284 |
0,000 |
|
0,716 |
|
u = 0 |
0,287 |
0,287 |
0,000 |
|
0,713 |
|
b = 0 |
0,286 |
0,286 |
0,000 |
|
0,714 |
|
h = z = 0 |
0,224 |
0,000 |
0,224 |
|
0,776 |
|
c = y = 0, i = v = 0 |
0,250 |
0,250 |
0,000 |
|
0,750 |
Tabla 7. Valores de la varianza transmisible y la varianza ambiental no transmisible para el Indice de Quetelet bajo todos los modelos de transmisión contrastados.
Table 7. Values of the transmissible and the non-transmissible variance for the Quetelet9s Index under all the tested models of transmission.
Los estudios que han tratado de cuantificar los componentes genéticos y ambientales, que actúan en la transmisión familiar de los rasgos de herencia multifactorial, no son demasiado abundantes en la literatura científica (Rao et al. 1982, 1984, Byard et al. 1985 Devor y Crawford 1984, Pérusse et al. 1988). Esto se ha debido en muchos casos a lo difícil que resulta conseguir un tamaño muestral adecuado cuando se trata de datos de individuos biológicamente emparentados, así como a la carencia de herramientas informáticas adecuadas para tratar la información. Además, los escasos trabajos en los que se ha empleado el método del análisis path para estudiar variables antropométricas han utilizado habitualmente el modelo TAU, frente al BETA empleado por nosotros. Esta escasez de investigaciones va a limitar enormemente la discusión de nuestros resultados. Sin embargo, no debemos pasar por alto que las estimas de los factores transmisibles son específicas para cada población o muestra poblacional (Pérusse et al. 1987).
El hecho de que el modelo general de transmisión familiar, estimador de 10 parámetros, se haya ajustado satisfactoriamente a nuestros datos, nos permite afirmar que puede tratarse del modelo menos restrictivo que definiría mejor la transmisión del índice de Quetelet. Asimismo, el hecho de que el modelo de transmisión sin efectos genéticos no haya mostrado un buen ajuste pone de relieve la importancia del componente hereditario sobre la transmisión de esta variable dentro de las familias. Sin embargo, debe existir un componente no genético que, en mayor o menor medida, esté actuando sobre la determinación del fenotipo del índice de Quetelet, dado que el modelo de transmisión sin herencia cultural no se ha ajustado bien para esta variable. Tampoco fue posible aceptar la hipótesis de una transmisión familiar sin homogamia social entre los cónyuges. Esto podría atribuirse al efecto de la convivencia tras el matrimonio sobre la semejanza entre los esposos. Incluso en una muestra socioeconómicamente homogénea, como la analizada, el modelo mostró que la homogamia está claramente presente. Byard et al. (1985) han señalado que la semejanza familiar observada para el peso en una muestra hindú, sería debida fundamentalmente a la homogamia social y a la cohabitación, pues la dieta y el estilo de vida comunes son factores determinantes en el peso después del matrimonio. Estos factores también han sido señalados por Province y Rao (1985) en su estudio sobre la transmisión familiar para la estatura, el peso y el IMC en una muestra de Brasil.
En la literatura sobre genética cuantitativa humana es muy frecuente encontrar alusiones a que tal o cual rasgo es altamente familiar, y que se transmite de padres a hijos con un componente cultural que debe ser importante. Bouchard (1996), en un estudio sobre la genética de la obesidad humana, ha señalado que hay al menos 4 loci ligados con el índice de masa corporal, y estudios realizados en gemelos idénticos (MZ) han demostrado la estrecha relación entre el IMC de los niños y sus padres biológicos, pero no con el de sus padres adoptivos (Stunkard et al. 1990), habiéndose sugerido incluso que los factores hereditarios serían responsables de hasta el 70% de las diferencias en el índice de Quetelet de individuos adultos (Sims 1990). El grado de determinación familiar, referida a la transmisión cultural de padres a hijos, viene dado en nuestro análisis por los parámetros fP y fM. En base a los valores estimados para estos dos parámetros en la muestra vasca, podemos entrever una posible existencia de efectos maternos sobre el índice estudiado. Esto viene, asimismo, apoyado por las mayores correlaciones entre los fenotipos de la madre y del padre con el fenotipo de los hijos (Tabla 5). En un reciente trabajo realizado en una muestra de esta misma zona del País Vasco (Salces et al. 2001a), se han encontrado evidencias de un posible efecto materno para la semejanza familiar del IMC. Tras dividir la muestra familiar total según si la madre era ama de casa o tenía un trabajo fuera de ésta, se siguió detectando una mayor semejanza de los hijos con la madre para el índice de Quetelet. Es decir, que el tiempo de permanencia de la madre en el hogar se exteriorizaba como una mayor semejanza familiar de ésta con sus descendientes, tomando como valor control la semejanza de los hijos con el padre. Province y Rao (1985) han observado, de igual forma, evidencias de un efecto materno que estaría actuando sobre la transmisión del peso en familias brasileñas, pero no llegaron a la misma conclusión para la estatura y el IMC, al contrario de lo observado en la muestra vasca, mientras que Pérusse et al. (1988) han encontrado transmisibilidades similares en ambos progenitores para el IMC.
Los estudios publicados sobre la transmisión familiar del índice de Quetelet en familias nucleares, bajo un modelo BETA de análisis path, son muy escasos. Sin embargo, los existentes han señalado componentes transmisibles explicativos del 35-40% de la varianza fenotípica observada (Bouchard 1985, Bouchard et al. 1988, Pérusse et al. 1988), superiores en todos los casos, al valor estimado para la muestra vasca (t2 = 28%).
En conclusión, el modelo que mejor ajusta la transmisión del índice de Quetelet dentro de las familias analizadas ha sido el de 10 parámetros (modelo general). Este es, probablemente, el modelo que mejor se ajusta a los datos estudiados. El análisis de los componentes transmisibles (genéticos y culturales) así como el de los factores ambientales no transmisibles sobre la semejanza familiar, ha mostrado evidencia de efectos tanto genéticos como no genéticos que podrían estar actuando sobre la determinación del fenotipo del IMC. Las mayores correlaciones observadas entre los hijos y la madre con respecto a las existentes entre los hijos y el padre, así como la mayor magnitud relativa del parámetro fM, muestran una mayor influencia materna que paterna sobre la herencia del índice de masa corporal. Las estimas del componente transmisible de la varianza en la muestra vasca han sido más bajas que las encontradas en la literatura que ha empleado el modelo BETA de análisis path. Sin embargo, no debemos olvidar que estas estimas son específicas para cada población.
Agradecimientos. Esta investigación ha sido parcialmente financiada por un proyecto de investigación de la Universidad del País Vasco (UPV154.310-EA049/99) y por el Gobierno Vasco (Departamento de Educación, Universidades e Investigación) a través de una beca predoctoral así como por una ayuda para que I. Salces realizase una estancia en las Universidades Libres de Bruselas (U.L.B. y V.U.B.).
Bouchard, C., 1985, Inheritance of fat distribution and adipose tissue metabolism. En Metabolic complications of human obesities, editado por J. Vague, P. Björntorp, B. Guy-Grand, M., Rebuffé-Scrive y P. Vague (Elsevier: Amsterdam), 87-96.
Bouchard, C., 1996, The genetics of obesity in humans. Current Opinion in Endocrinology and Diabetes, 3, 29-35.
Bouchard, C., Pérusse, L., Leblanc, C., Tremblay, A., y Thériault, G., 1988, Inheritance of the amount and distribution of human body fat. International Journal of Obesity, 12, 205-215.
Byard, P.J., Poosha, D.V.R., y Satyanarayana, M., 1985, Genetic and environmental determinants of height and weight in families from Andhra Pradesh, India. Human Biology, 57, 621-633.
Cloninger, C.R., Rice, J., y Reich, T., 1979a, Multifactorial inheritance with cultural transmission and assortative mating. II. A general model of combined polygenic and cultural inheritance. American Journal of Human Genetics, 31, 176-198.
Cloninger, C.R., Rice, J., y Reich, T., 1979b, Multifactorial inheritance with cultural transmission and assortative mating. III. Family structure and the analysis of separation experiments. American Journal of Human Genetics, 31, 366-388.
Devor, E.J., y Crawford, M.H., 1984, Family resemblance for neuromuscular performance in a Kansa Mennonite community. American Journal of Physical Anthropology, 67, 289-296.
Longini, I.M., M.W. Higgins, P.C., Hinton, P.P. Moll, y J.B. Keller, 1984, Genetic and environmental sources of familial aggregation of body mass in Tecumseh, Michigan. Human Biology, 56, 733-757.
Mueller, W.H., 1983, The genetics of human fatness. Yearbook of Physical Anthropology, 26, 215-230.
Mueller, W.H., y Malina, R.M., 1980, Genetic and environmental influences on growth of Philadelphia black and white schoolchildren. Annals of Human Biology, 7, 441-448.
OMS, 1995, El estado físico: uso e interpretación de la antropometría. Serie de informes técnicos, nº 854 (Ginebra: Organización Mundial de la Salud).
Pérusse, L., Leblanc, C., y Bouchard, C., 1988, Inter-generation transmission of physical fitness in the Canadian population. Canadian Journal of Sportif Sciences, 13, 8-14.
Pérusse, L., Lortie, G., Leblanc, C., Tremblay, A., Thériault, G., y Bouchard, C., 1987, Genetic and environmental sources of variation in physical fitness. Annals of Human Biology, 14, 425-434.
Province, M.A., y Rao., D.C., 1985, Path analysis of family resemblance with temporal trends: Applications to height, weight, and Quetelet index in Northeastern Brazil. American Journal of Human Genetics, 37, 178-192.
Rao, D.C., Laskarzewski, P.M., Morrison, J.A., Khoury, P., Kelly, K., Wette, R., Russell, J., y Glueck, C.J., 1982, The Cincinnati Lipid Research Clinic Family Study: Cultural and Biological Determinants of Lipids and Lipoprotein Concentrations. American Journal of Human Genetics, 34, 888-903.
Rao, D.C., Maclean, C.J., y Morton, N.E., 1975, Analysis of family resemblance. V. Height and weight in Northeastern Brazil. American Journal of Human Genetics, 27, 509-520.
Rao, D.C., McGue, M., Wette, R., y Glueck, C.J., 1984, Path analysis in genetic epidemiology. En Human population genetics: the Pittsburgh Symposium, editado por A. Chakravarti. (New York: Van Nostrand Reinhold Company), 35-81.
Rao, D.C., Morton, N.E., y Cloninger, C.R., 1979, Path analysis under generalized assortative mating. Genetic Research, Cambridge, 33, 175-188.
Rebato, E., Salces, I., San Martín, L., Rosique, J., Hauspie, R., y Susanne, C., 1997, Age variations in sibling correlations for height, sitting height and weight. Annals of Human Biology, 24, 58-592.
Rice, J., Cloninger, C.R., y Reich, T., 1978, Multifactorial inheritance with cultural transmission and assortative mating. I. Description and basic properties of the unitary models. American Journal of Human Genetics, 30, 618-643.
Roberts, D.F., 1985, Genetic contribution to phenotype variation in growth and body composition. Human Biology Budapest, 16, 618-643.
Salces, I., Rebato, E., San Martín, L., Rosique, J., Vinagre, A., y Susanne, C., 2001a, Family resemblance for anthropometric traits. II. Assessment of occupational maternal and age effects. HOMO (en revisión)
Salces, I., Rebato, E., Susanne, C., San Martín, L., Rosique, J., y Vinagre, A., 2001a, Family resemblance for anthropometric traits. I. Evaluation of maternal occupation influence. Biométrie Humaine et Anthropologie (en prensa).
Sánchez Andrés, A., y Mesa, M.S., 1994, Heritabilities of morfological and body composition characteristics in a Spanish population. Anthropologischer Anzeiger, 52, 341-349.
Sims, E.A.H., 1990,Destiny rides again as twins overeat. New England Journal of Medicine, 322, 1522-1524.
Sørensen, T.I.A., Price, R.A., Stunkard, A.J., y Schulsinger, F., 1989, Genetics of obesity in adult adoptees and their biological siblings. British Medical Journal, 298, 87-90.
Stunkard, A.J., Harris, J.R., Pedersen, N.L., McClearn, G.E., 1990, The body-mass index of twins who have been reared apart. New England Journal of Medicine, 322, 1483-1487.
Susanne, C., 1974, Les changements morphologiques liés à l'âge. L'Anthropologie, 78, 693-718.
Susanne, C., 1977, Individual age changes of the morphological characteristics. Journal of Human Evolution, 6, 181-189.
Weiner, J.S., y Lourie, J.A., 1981, Practical Human Biology (London: Academic Press).
Wright, S., 1921, Correlation and causation. Journal of Agricultural Research, XX, 557-585.