
Peña,
J. A., 2001. Estructura genética de la población del Valle de Orozco (País
Vasco, España) a partir de diferentes matrices de migración. Antropo, 0,
1-10. www.didac.ehu.es/antropo
Estructura genética de la población del Valle de Orozco (País Vasco, España) a partir de diferentes matrices de migración
Dpto. Biología Animal y Genética. Facultad de Ciencias. Universidad del País Vasco. Aptdo. 644. 48080 Bilbao (SPAIN). Tel: 946 012 600. E-mail: ggppegaj@lg.ehu.es
Palabras
clave: Estructura genética,
matrices de migración, ![]()
Se ha comparado la capacidad de varios tipos de matrices de migración para describir
el flujo génico. Las matrices empleadas han sido: 1) progenitores-descendientes,
2) padres-descendientes, 3) madres-descendientes, 4) progenitores-descendientes con
reconstrucción de genealogías, 5) domicilio de los progenitores-origen
de los descendientes con reconstrucción de genealogías, 6) origen de los
cónyuges y 7) domicilio de los cónyuges. Utilizando el método descrito
por Imaizumi et al (1970) y Harpending y Jenkins (1973), se ha proyectado el patrimonio
genético de la población a una situación de equilibrio entre flujo
génico y deriva. Mediante esta simulación se ha obtenido una medida de
la varianza estandarizada de las frecuencias génicas (![]() )
y del parentesco entre las subpoblaciones.
)
y del parentesco entre las subpoblaciones.
Se han detectado diferencias entre las matrices. Las tasas de microdiferenciación obtenidas a partir de las matrices domicilio del varón-domicilio de la mujer y residencia de los progenitores-origen de los descendientes con reconstrucción de genealogías son mayores que las tasas obtenidas a partir de la matriz progenitores-descendientes, mostrando que la mayor parte del flujo génico tiene su origen en desplazamientos que ocurren entre la parroquia de nacimiento y la parroquia de residencia premarital de los individuos. No obstante, la topología del parentesco es descrita adecuadamente tan sólo a partir de las matrices intergeneracionales.
The ability for describing the gene flow from several kinds of migration matrices
has been compared. The used matrices are: 1) parent-offspring, 2) father-offspring,
3) mother-offspring, 4) parent-offspring with genealogical reconstruction, 5) parent's
residence-offspring's residence with genealogical reconstruction, 6) husband's origin-wife's
origin, and 7) husband's residence-wife's residence. Using procedure described by
Imaizumi et al (1970) and
Harpending and Jenkins (1973), the gene pool of the population has been projected
to the equilibrium between gene flow and genetic drift. With this simulation, the
standardized variance of gene frequencies, ![]() ,
and kinship coefficients between subpopulations are obtained.
,
and kinship coefficients between subpopulations are obtained.
Differences between matrices are detected. Microdifferentiation rates obtained from husband's residence-wife's residence and parent's residence-offspring's residence with genealogical reconstruction matrices are greater than the rates obtained from parent-offspring matrix, revealing that most of the gene flow has its origin in movements happening between the parish of birth and the parish of premarital residence of the individuals. However, topology of kinship is adequately described only from inter-generational matrices.
Las matrices de migración describen los patrones de movilidad de una población. Cada individuo puede desplazarse entre localidades en diferentes ocasiones a lo largo de su vida, pero no todos los movimientos tendrán trascendencia para la estructura de la población; la tendrán aquellos que se encuentren asociados a determinados hitos de su existencia (nacimiento, matrimonio, nacimiento de su descendencia). Por un lado, serán determinantes para el flujo génico los vectores resultantes de los desplazamientos entre el lugar de nacimiento de un individuo y el lugar de nacimiento de sus descendientes. Por otro, los desplazamientos desde el lugar de nacimiento o residencia hasta el lugar de matrimonio, definirán los patrones de búsqueda del cónyuge.
En función de los acontecimientos vitales que se utilicen para construir una matriz de migración, se estarán considerando distintos aspectos de la movilidad de los individuos de una población. En todo caso, no siempre será posible disponer de la matriz de migración adecuada para un determinado estudio, dependiendo de la calidad y la fuente de los datos, por lo que será preciso conocer en qué medida se pueden considerar componentes de la migración diferentes de los deseados.
Para estimar el parentesco entre poblaciones mediante modelos basados en matrices
de migración deberían utilizarse matrices progenitores-descendientes (lugar
de nacimiento de los progenitores frente a lugar de nacimiento de los descendientes).
No obstante, hay una serie de antecedentes en los que han sido utilizadas además
otras matrices con motivos comparativos. Entre otros, Workman et al.
(1973), han comparado la matriz progenitores-descendientes con la matriz lugar de
nacimiento-lugar de residencia, para los indios Papago, encontrando una concordancia
sustancial en los resultados. Jorde et al.
(1982) han utilizado siete tipos de matrices para las Islas Åland: progenitores-descendientes,
padres-descendientes, madres-descendientes, migración marital, origen-residencia,
origen-residencia de los varones y origen-residencia de las mujeres; han estimado
![]() y la correlación entre el
parentesco y la distancia geográfica para cada caso, encontrando unos patrones
similares. Roberts (1982) ha obtenido valores de
y la correlación entre el
parentesco y la distancia geográfica para cada caso, encontrando unos patrones
similares. Roberts (1982) ha obtenido valores de ![]() prácticamente idénticos y coeficientes de parentesco similares, a partir
de las matrices progenitores-descendientes, padres-descendientes y madres-descendientes
para la población de Cumbria. Mielke et al.
(1982) han hallado patrones similares de asociación entre pueblos en las Islas
Åland a partir de las matrices lugar de residencia pre y postmarital, origen
de los cónyuges y progenitores-descendientes. Jorde (1984), por último,
no ha observado grandes diferencias entre la matriz progenitores-descendientes y
la matriz origen de los progenitores-origen del hijo primogénito en Utah, concluyendo
que la segunda supone una medida correcta del flujo génico en una población
con una gran movilidad y una considerable fertilidad diferencial.
prácticamente idénticos y coeficientes de parentesco similares, a partir
de las matrices progenitores-descendientes, padres-descendientes y madres-descendientes
para la población de Cumbria. Mielke et al.
(1982) han hallado patrones similares de asociación entre pueblos en las Islas
Åland a partir de las matrices lugar de residencia pre y postmarital, origen
de los cónyuges y progenitores-descendientes. Jorde (1984), por último,
no ha observado grandes diferencias entre la matriz progenitores-descendientes y
la matriz origen de los progenitores-origen del hijo primogénito en Utah, concluyendo
que la segunda supone una medida correcta del flujo génico en una población
con una gran movilidad y una considerable fertilidad diferencial.
En este trabajo se analizan una serie de matrices de migración de la población del Valle de Orozco. Las diferencias en la medida de la variabilidad y el parentesco obtenidas por cada una de ellas revelan aspectos de interés de los diversos componentes de la movilidad de su población.
El Valle de Orozco es un municipio de la provincia de Vizcaya (País Vasco), situado entre 43º 2' 8'' y 43º 8' 45'' de latitud Norte y entre 2º 47' 17'' y 2º 58' 25'' de longitud Oeste, con una superficie de 102,4 Km2. Presenta una disposición longitudinal, según un eje Sudeste-Noroeste, de modo que la zona Sudeste es más montañosa que la zona Noroeste, más llana y abierta hacia el exterior (figura 1).
La población del valle ha sufrido diferentes altibajos desde principios del siglo XVIII, oscilando entre 2.000 y 3.000 habitantes. Una parte se encuentra dispersa por toda la superficie habitable, en pequeñas agrupaciones de caseríos. El resto se aglomera en cuatro núcleos principales, que se corresponden con las cuatro parroquias en que se divide:
1. Murueta, se encuentra situada en la zona más baja y mejor comunicada con el exterior y comprende aproximadamente el 12% de la población.
2. Zubiaur, es el núcleo principal del valle, con un 49% de la población.
3. Albizu, se halla en la zona ascendente y supone el 16% de los habitantes del valle.
4. Olarte, es la parroquia más elevada y agrupa al 23% de los individuos.
Se han recogido las actas parroquiales de los 4.798 matrimonios celebrados en el Valle de Orozco entre 1710 y 1984, en las que se encuentran registrados los lugares de nacimiento y de residencia de los cónyuges (en el 84% y el 69% de los casos respectivamente) y los lugares de nacimiento de sus progenitores (en el 72% de los casos). Con posterioridad, se ha realizado una reconstrucción de genealogías, obteniéndose de nuevo los lugares de nacimiento de los progenitores, así como sus lugares de residencia premarital (en el 41% de los casos).
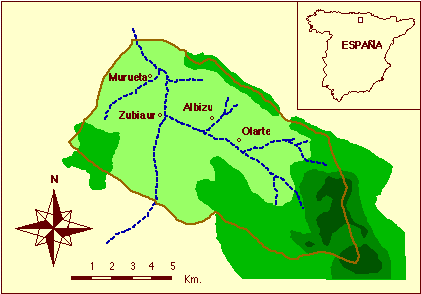
Figura 1. El Valle de Orozco, provincia de Vizcaya (País Vasco, España).
Figure 1. The Orozco Valley, Biscay province (Basque Country, Spain).
Con estos datos, se han construido las siguientes matrices de migración:
1. Progenitores-descendientes. Se consideran los lugares de nacimiento de los progenitores y de sus descendientes. Cada progenitor se encuentra representado en la matriz tantas veces como hijos suyos hayan nacido en el valle y cada descendiente dos veces, una por su padre y otra por su madre.
2. Padres-descendientes.
3. Madres-descendientes.
4. Progenitores-descendientes con reconstrucción de genealogías.
5. Domicilio de los progenitores-origen de los descendientes, con reconstrucción de genealogías.
6. Origen de los cónyuges. Se define como el lugar de nacimiento del marido frente al lugar de nacimiento de la esposa.
7. Domicilio de los cónyuges, construida a partir de los lugares de residencia premarital de ambos cónyuges.
Las cinco primeras son matrices intergeneracionales, por lo que deben describir en mayor o menor medida el flujo génico; las dos últimas, por el contrario son matrices de migración marital, de modo que definen los patrones de búsqueda del cónyuge.
En cada caso se ha calculado la matriz estocástica, P, cuyos elementos ![]() definen la probabilidad de que un gen de la subpoblación j provenga de la subpoblación
i.
definen la probabilidad de que un gen de la subpoblación j provenga de la subpoblación
i.
Se han estimado los tamaños efectivos de cada subpoblación como un tercio del tamaño medio de cada parroquia a lo largo del período estudiado. Partiendo de P y Ne, se han obtenido las matrices de parentesco a priori, según el método de Malécot (1950), desarrollado por Imaizumi et al. (1970), que puede describirse como sigue:
![]()
donde ![]() es el vector de valores
de presión sistemática,
es el vector de valores
de presión sistemática,
P es la matriz de migración estocástica, y
![]() es el tamaño efectivo de
la subdivisión k.
es el tamaño efectivo de
la subdivisión k.
A continuación, se ha transformado la matriz de parentesco a priori,
![]() , en una matriz de parentesco
condicional, R, según el método de Harpending y Jenkins (1973):
, en una matriz de parentesco
condicional, R, según el método de Harpending y Jenkins (1973):
![]()
donde
![]()
![]() es la proporción de la población
total que vive en i, y
es la proporción de la población
total que vive en i, y
![]()
La matriz R especifica el parentesco relativo a una serie de frecuencias de genes
contemporánea, en tanto que la matriz ![]() lo predice en relación a una serie ancestral. En ambos casos, los elementos
de la matriz obtenida especifican la probabilidad de que un gen dado tomado de una
población i, sea idéntico por descendencia a otro gen homólogo tomado
de otra población j.
lo predice en relación a una serie ancestral. En ambos casos, los elementos
de la matriz obtenida especifican la probabilidad de que un gen dado tomado de una
población i, sea idéntico por descendencia a otro gen homólogo tomado
de otra población j.
Además, se ha obtenido una medida de la variabilidad entre las poblaciones,
mediante la fórmula ![]() , que es
una estima directa del estadístico
, que es
una estima directa del estadístico ![]() de Wright. Puesto que
de Wright. Puesto que ![]() se ve
influído por el tamaño de la población y el número de subdivisiones,
para su interpretación se ha multiplicado por -1 el logaritmo de la probabilidad
del producto
se ve
influído por el tamaño de la población y el número de subdivisiones,
para su interpretación se ha multiplicado por -1 el logaritmo de la probabilidad
del producto ![]() en la
función
en la
función ![]() , donde
, donde
![]() es el tamaño efectivo de
la población y r es el número de subdivisiones. -log(p) es una función
lineal de la heterogeneidad, que no se ve influído por el tamaño muestral
(Peña et al., 1997).
es el tamaño efectivo de
la población y r es el número de subdivisiones. -log(p) es una función
lineal de la heterogeneidad, que no se ve influído por el tamaño muestral
(Peña et al., 1997).
Para la representación en un espacio euclídeo de las matrices de parentesco se ha utilizado el análisis de escalamiento multidimensional y el análisis de escalamiento individual. En ambos casos se requieren matrices de disimilaridad, por lo que los coeficientes de parentesco han sido transformados en coeficientes de disimilaridad mediante la fórmula:
![]()
cuyo resultado cumple las propiedades de la distancia euclídea (Crawford y Mielke, 1982; Mielke et al, 1982; Roberts, 1982).
El análisis de escalamiento individual permite obtener una solución de consenso cuando se tienen en cuenta varias matrices referidas a un mismo grupo de subpoblaciones (Carroll y Chang, 1970; Gower, 1975; Cuadras, 1981). El análisis ofrece una representación conjunta de las subpoblaciones mediante unas coordenadas en un número de dimensiones reducido, pero además permite obtener una representación de las matrices e incluso una representación diferente de las subpoblaciones a partir de cada matriz.
A partir de las diferentes matrices de migración, se han estimado las matrices R de parentesco entre las parroquias del Valle de Orozco. El número de iteraciones necesario para alcanzar la situación de equilibrio ha sido muy variable: si bien en algunos casos se han necesitado hasta 80 (matriz de domicilios de los cónyuges), para la matriz progenitores-descendientes ha bastado con 15. No obstante, no pueden identificarse iteraciones con generaciones. De hecho, aplicando el modelo de Bodmer y Cavalli-Sforza (1968), fueron necesarias únicamente 5 iteraciones para la misma matriz progenitores-descendientes.
Los valores de ![]() obtenidos
para cada matriz se muestran en la figura 2. Como se observa, oscilan entre 0,00077
para la matriz madres-descendientes y 0,00443 para la matriz de domicilios de los
cónyuges.
obtenidos
para cada matriz se muestran en la figura 2. Como se observa, oscilan entre 0,00077
para la matriz madres-descendientes y 0,00443 para la matriz de domicilios de los
cónyuges.
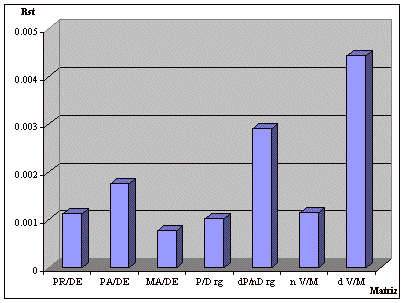
Figura
2. Valores
de ![]() obtenidos a partir
de diferentes matrices de migración. PR/DE: progenitores-descendientes; PA/DE:
padres-descendientes; MA/DE: madres-descendientes; P/D rg: progenitores-descendientes
con reconstrucción de genealogías; dP/nD rg: domicilio de los progenitores-origen
de los descendientes con reconstrucción de genealogías; n V/M: origen de
los cónyuges; d V/M: domicilio de los cónyuges.
obtenidos a partir
de diferentes matrices de migración. PR/DE: progenitores-descendientes; PA/DE:
padres-descendientes; MA/DE: madres-descendientes; P/D rg: progenitores-descendientes
con reconstrucción de genealogías; dP/nD rg: domicilio de los progenitores-origen
de los descendientes con reconstrucción de genealogías; n V/M: origen de
los cónyuges; d V/M: domicilio de los cónyuges.
Figure
2. ![]() values obtained from different migration matrices. PR/DE: parent-offspring; PA/DE:
father-offspring; MA/DE: mother-offspring; P/D rg: parent-offspring with genealogical
reconstruction; dP/nD rg: parent's residence-offspring's residence with genealogical
reconstruction; n V/M: husband's origin-wife's origin; d V/M: husband's residence-wife's
residence.
values obtained from different migration matrices. PR/DE: parent-offspring; PA/DE:
father-offspring; MA/DE: mother-offspring; P/D rg: parent-offspring with genealogical
reconstruction; dP/nD rg: parent's residence-offspring's residence with genealogical
reconstruction; n V/M: husband's origin-wife's origin; d V/M: husband's residence-wife's
residence.
La matriz más adecuada para predecir el equilibrio migración-deriva es
la que conforman los lugares de nacimiento de los progenitores y de sus descendientes.
En este caso, ![]() toma un
valor de 0,00112, que se corresponde con un valor de -log(p) de 0,24. Por tanto,
no se espera una heterogeneidad genética significativa en la población
del Valle de Orozco, ya que -log(p) debería ser mayor de 1,3 para una probabilidad
de 0,05 (Peña et al.,
1997).
toma un
valor de 0,00112, que se corresponde con un valor de -log(p) de 0,24. Por tanto,
no se espera una heterogeneidad genética significativa en la población
del Valle de Orozco, ya que -log(p) debería ser mayor de 1,3 para una probabilidad
de 0,05 (Peña et al.,
1997).
El parámetro ![]() obtenido
a partir de matrices de migración muestra la influencia que sobre la variabilidad
de la población presentan los patrones de migración; por tanto, da una
idea de la magnitud de la migración con respecto al tamaño efectivo de
la población. Puesto que el tamaño efectivo es el mismo en todos los casos,
obtenido
a partir de matrices de migración muestra la influencia que sobre la variabilidad
de la población presentan los patrones de migración; por tanto, da una
idea de la magnitud de la migración con respecto al tamaño efectivo de
la población. Puesto que el tamaño efectivo es el mismo en todos los casos,
![]() podrá tomarse como una medida
de la movilidad descrita por las diferentes matrices de migración, de modo que,
básicamente, una mayor varianza implicará una menor inmigración (Jorde,
1984).
podrá tomarse como una medida
de la movilidad descrita por las diferentes matrices de migración, de modo que,
básicamente, una mayor varianza implicará una menor inmigración (Jorde,
1984).
Los valores resultantes muestran algunos aspectos de interés:
a) Las matrices progenitores-descendientes, obtenidas a partir de los registros de matrimonios y de la reconstrucción de genealogías, presentan unos niveles muy similares (0,00112 y 0,00103, para unos valores de -log(p) de 0,24 y 0,21 respectivamente).
b) La matriz padres-descendientes predice un mayor valor que la matriz madres-descendientes (0,00175 y 0,00077, para unos valores de -log(p) de 0,42 y 0,14 respectivamente), por lo que las mujeres presentan una mayor movilidad. Es decir, se detecta una marcada patrilocalidad, de modo que los varones se desplazan en menor medida que las mujeres entre su lugar de nacimiento y el lugar en el que tienen su descendencia. De hecho, en tanto que el 70,25% de los individuos que contraen matrimonio en el Valle de Orozco son hijos de padres varones nacidos en Orozco, sólo el 66,29% son hijos de madres nacidas en este mismo lugar. Este mismo hecho ha sido detectado por Cavalli-Sforza y Bodmer (1971), Mielke et al. (1982) y Jorde et al. (1982) en otras poblaciones.
c) La matriz domicilio de los progenitores-origen de los descendientes predice valores
más altos de ![]() que el
resto de matrices intergeneracionales (0,00291, para un valor de -log(p) de 0,78),
lo que implica una menor movilidad. Esta matriz describe una parte del flujo génico,
específicamente aquella que se da entre el lugar de residencia de los progenitores
y el lugar de nacimiento de sus descendientes. Las diferencias con la matriz progenitores-descendientes
vendrán determinadas por los desplazamientos realizados por los individuos entre
su lugar de nacimiento y su lugar de residencia.
que el
resto de matrices intergeneracionales (0,00291, para un valor de -log(p) de 0,78),
lo que implica una menor movilidad. Esta matriz describe una parte del flujo génico,
específicamente aquella que se da entre el lugar de residencia de los progenitores
y el lugar de nacimiento de sus descendientes. Las diferencias con la matriz progenitores-descendientes
vendrán determinadas por los desplazamientos realizados por los individuos entre
su lugar de nacimiento y su lugar de residencia.
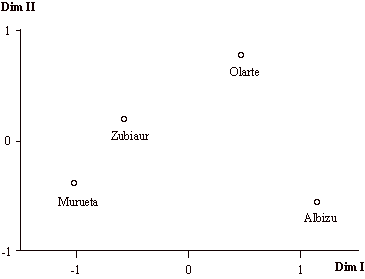
Figura 3. Análisis de escalamiento multidimensional de las distancias obtenidas a partir de la matriz progenitores-descendientes para las parroquias del Valle de Orozco.
Figure 3. Multidimensional scaling analysis of the distance matrix obtained from the parent-offspring migration matrix for the parishes of the Orozco Valley.
d) Las matrices origen de los cónyuges y domicilio de los cónyuges describen
otro tipo de movimientos, concretamente la migración marital, por lo que a
priori no son comparables
con las matrices intergeneracionales. No obstante, la primera produce valores similares
a la matriz progenitores-descendientes (0,00114; -log(p)=0,24), en tanto que la segunda
sugiere una movilidad mucho menor (![]() =0,00443;
-log(p)=1,29). La diferencia entre ambas matrices vendrá dada, del mismo modo
que en el caso anterior, por la movilidad entre el lugar de nacimiento y el lugar
de residencia premarital de cada cónyuge.
=0,00443;
-log(p)=1,29). La diferencia entre ambas matrices vendrá dada, del mismo modo
que en el caso anterior, por la movilidad entre el lugar de nacimiento y el lugar
de residencia premarital de cada cónyuge.
Si bien el parámetro ![]() puede
informar sobre la magnitud de la migración, no define su distribución entre
las subpoblaciones, de modo que partiendo de dos matrices muy diferentes podrían
obtenerse valores de varianza idénticos. Por lo tanto, es interesante considerar
también el grado de parentesco entre cada una de las parroquias del Valle de
Orozco.
puede
informar sobre la magnitud de la migración, no define su distribución entre
las subpoblaciones, de modo que partiendo de dos matrices muy diferentes podrían
obtenerse valores de varianza idénticos. Por lo tanto, es interesante considerar
también el grado de parentesco entre cada una de las parroquias del Valle de
Orozco.
En primer lugar se han calculado los coeficientes de parentesco condicional a partir de la matriz progenitores-descendientes. Una vez transformados en disimilaridades, han dado lugar a la representación de la figura 3 mediante un análisis de escalamiento multidimensional. El estrés de la configuración final después de 12 iteraciones es 0, por lo que el ajuste puede considerarse perfecto.
La distribución de las parroquias respecto a las dos dimensiones obtenidas no se corresponde exactamente con su posición geográfica. La principal diferencia se encuentra en Albizu, población situada en una zona intermedia del valle que, sin embargo, aparece poco emparentada con el resto de las parroquias. Según las dos dimensiones obtenidas, se sitúan a una máxima distancia las poblaciones de Murueta y Albizu, de modo que serán las poblaciones menos emparentadas, tanto entre ellas como con el conjunto de las parroquias de Orozco.
El coeficiente de aislamiento reproductor (Lasker y Kaplan, 1964) puede ayudar a la interpretación de estos resultados. Dicho coeficiente se define como el producto del tamaño efectivo de población y la tasa de inmigración efectiva, de modo que los valores más pequeños indicarán una mayor posibilidad de aislamiento y acción de la deriva (Jorde, 1982). Como se observa en la tabla 1, los coeficientes más bajos corresponden a Albizu y Murueta, por lo que se espera que sean estas dos parroquias las que tengan un flujo génico menor y por tanto se encuentren menos relacionadas con el resto de parroquias.
|
Parroquia |
|
|
Murueta |
18,9 |
|
Zubiaur |
58,7 |
|
Albizu |
14,6 |
|
Olarte |
30,4 |
Tabla 1. Coeficiente de aislamiento reproductor en las parroquias del Valle de Orozco.
Table 1. Coefficient of breeding isolation for the parishes of the Orozco Valley.
En un siguiente paso, se ha realizado un análisis de escalamiento individual considerando conjuntamente todas las matrices. Después de 5 iteraciones, el estrés global resultante ha sido del 9%, por lo que el ajuste puede considerarse bueno. La varianza total explicada es el 92,5%.
La ubicación topológica de las subpoblaciones respecto a las dos dimensiones obtenidas es una solución de compromiso, altamente influenciada por la introducción en el análisis de matrices no adecuadas. Como puede observarse en la figura 4, las diferencias con la configuración obtenida exclusivamente a partir de la matriz progenitores-descendientes (figura 3) son sustanciales. Si bien continúan siendo las parroquias de Murueta y Albizu las menos emparentadas, ahora Zubiaur muestra un menor parentesco con el resto del valle en tanto que Olarte, por el contrario, pasa a ser el núcleo más relacionado con el resto de subpoblaciones.
El aspecto más significativo de este análisis radica en la posibilidad de establecer comparaciones entre las diferentes matrices de migración. En la figura 5 se representan los pesos de cada una de las matrices respecto a las dos dimensiones obtenidas en la figura 4. Los principales resultados que pueden destacarse son los siguientes:
a. La matrices progenitores-descendientes, padres-descendientes y madres-descendientes muestran una idéntica relación de parentesco entre las parroquias del Valle de Orozco, lo que implica unos patrones de migración muy similares.
b. La matriz progenitores-descendientes con reconstrucción de genealogías representa una notable aproximación al modelo del resto de matrices intergeneracionales.
c. Las matrices origen de los cónyuges y domicilio de los cónyuges presentan muy pocas similitudes con la matriz progenitores-descendientes; mientras que las primeras se ven en su mayor parte explicadas por la dimensión II, la segunda se interpreta exclusivamente por la dimensión I, por lo que recogerán aspectos notablemente diferentes de la movilidad de la población.
d. La matriz domicilio de los progenitores-origen de los descendientes se encuentra en una situación intermedia entre las matrices intergeneracionales y las matrices de migración marital, de modo que un 68% de su varianza se encuentra explicada por la dimensión II y un 24% por la dimensión I.
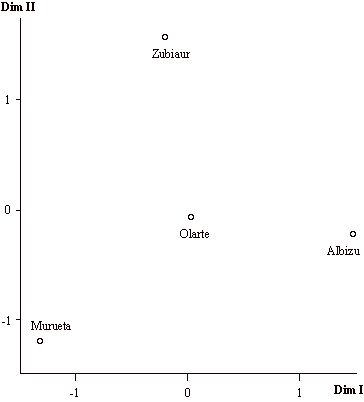
Figura 4. Análisis de escalamiento individual de las distancias obtenidas a partir de 7 matrices de migración para las parroquias del Valle de Orozco.
Figure 4. Individual scaling of the distance matrices obtained from 7 migration matrices for the parishes of the Orozco Valley.
La mayor parte de los desplazamientos que finalmente afectan al flujo génico y por tanto al patrimonio genético de la población del Valle de Orozco son premaritales, implicando los cambios entre el lugar de nacimiento y el lugar de residencia antes del matrimonio. Así, las matrices que asocian los lugares de nacimiento de ambos cónyuges predicen una heterogeneidad genética muy similar a las matrices progenitores-descendientes. Las variaciones debidas a patrilocalidad entre las matrices presentan una influencia menor. Estos resultado concuerdan con los recogidos en otros estudios, ya que se han obtenido similares valores entre las matrices progenitores-descendientes y lugar de nacimiento-lugar de residencia para los indios Papago (Workman et al., 1973), entre las matrices progenitores-descendientes, padres-descendientes, madres-descendientes, migración marital, origen-residencia, origen-residencia de los varones y origen-residencia de las mujeres para las Islas Åland (Jorde et al., 1982), entre las matrices progenitores-descendientes, padres-descendientes y madres-descendientes para la población de Cumbria (Roberts, 1982) y entre las matrices progenitores-descendientes y origen de los progenitores-origen del hijo primogénito en Utah (Jorde ,1984). Sin embargo, no concuerdan con los resultados descritos por Mielke et al. (1982) para las Islas Åland, donde observaron valores similares entre las matrices lugar de residencia pre y postmarital y progenitores-descendientes.
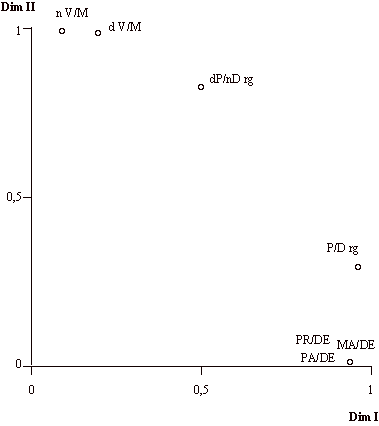
Figura 5. Pesos de las matrices en las dos dimensiones de la configuración global del análisis de escalamiento individual. PR/DE: progenitores-descendientes; PA/DE: padres-descendientes; MA/DE: madres-descendientes; P/D rg: progenitores-descendientes con reconstrucción de genealogías; dP/nD rg: domicilio de los progenitores-origen de los descendientes con reconstrucción de genealogías; n V/M: origen de los cónyuges; d V/M: domicilio de los cónyuges.
Figure 5. Weights of the matrices in the two dimensions of the global configuration of the individual scaling. PR/DE: parent-offspring; PA/DE: father-offspring; MA/DE: mother-offspring; P/D rg: parent-offspring with genealogical reconstruction; dP/nD rg: parent's residence-offspring's residence with genealogical reconstruction; n V/M: husband's origin-wife's origin; d V/M: husband's residence-wife's residence.
En Orozco, las matrices que asocian los lugares de residencia de ambos cónyuges o bien los lugares de residencia de los progenitores con los lugares de nacimiento de sus descendientes predicen una heterogeneidad mucho mayor y por tanto informan de una menor movilidad.
La mayor parte de los trabajos en los que se han comparado diferentes tipos de matrices
se basan en el cálculo de las tasas de microdiferenciación. Pero estas
tasas informan de la magnitud del flujo génico hacia una población y de
su interacción con la deriva genética, pero no de la migración entre
subdivisiones. Si se analiza la estructura genética de las subdivisiones, pueden
encontrarse diferencias importantes entre matrices que dan lugar a valores muy similares
de ![]() o
o ![]() .
En el Valle de Orozco se han encontrado diferencias cualitativas importantes en los
desplazamientos descritos por la matriz progenitores-descendientes en relación
a la matriz de migración marital.
.
En el Valle de Orozco se han encontrado diferencias cualitativas importantes en los
desplazamientos descritos por la matriz progenitores-descendientes en relación
a la matriz de migración marital.
Si la matriz progenitores-descendientes es la que describe más correctamente el flujo génico, ni las matrices de migración marital, ni la matriz domicilio de los progenitores-origen de los descendientes pueden considerarse idóneas para su estudio, pues aun cuando puedan mostrar una cantidad de variabilidad similar, la predicción de la estructura del parentesco varía considerablemente.
Agradecimientos. Este trabajo ha sido financiado parcialmente por el proyecto UPV 154.310-EA082/98 de la Universidad del País Vasco. Queremos mostrar nuestro agradecimiento al padre Beperet, al padre Ojanguren y al personal del Archivo Histórico Eclesiástico de Vizcaya por su colaboración en la recogida de datos.
Bodmer, W., y Cavalli-Sforza, L., 1968, A migration matrix model for the study of random genetic drift. Genetics, 59, 565-592.
Carroll, J. D., y Chang, J. J., 1970, Analysis of individual differences in multidimensional scaling via an N-way generalization of "Eckart-Young" Decomposition. Psychometrika, 35, 283-319.
Cavalli-Sforza, L. L., y Bodmer, W., 1971, Genética de las poblaciones humanas (Barcelona: Omega).
Crawford, M. y Mielke, J., 1982, Current developments in Anthropological Genetics. Volume 2: Ecology and population structure (New York: Plenum Press).
Cuadras, C., 1981, Métodos de análisis multivariante (Barcelona: Editorial Universitaria de Barcelona).
Gower, J., 1975, Generalized Procrustes analysis. Psychometrika, 40, 33-51.
Harpending, H., y Jenkins, T., 1973, Genetic distance among southern African populations. En Methods and theories of Anthropological Genetics, editado por M. H. Crawford y P. L. Workman (Albuquerque: University of New Mexico Press) pp. 177-199
Imaizumi, Y., Morton, N., y Harris, D., 1970, Isolation by distance in artificial populations. Genetics, 66, 569-582.
Jorde, L. B., Workman, P., y Eriksson, A., 1982, Genetic microevolution in the Åland Islands, Finland. En Current developments in Anthropological Genetics. Volume 2: Ecology and population structure, editado por M. Crawford y J. Mielke (New York: Plenum Press) pp. 333-365.
Jorde, L., 1982, The genetic structure of the Utah Mormons: migration analysis. Human Biology, 54, 583-597.
Jorde, L. B., 1984, A comparison of parent-offspring and marital migration data as measures of gene flow. En Migration and mobility, editado por A. J. Boyce (London: Taylor y Francis) pp. 83-96.
Lasker, G., y Kaplan, B., 1964, The coefficient of breeding isolation: Population size, migration rates, and the possibilities for random genetic drift in six human communities in Northern Peru. Human Biology, 36, 327-338.
Malécot, G., 1950, Quelques schémas probabilistes sur la variabilité des populations naturelles. Annales de l9Université de Lyon, Science Section A, 13, 37-60.
Mielke, J., Devor, E., Kramer, P., Workman, P., y Eriksson, A., 1982, Historical population structure of the Åland Islands, Finland. En Current developments in Anthropological Genetics. Volume 2: Ecology and population structure, editado por M. Crawford y J. Mielke (New York: Plenum Press) pp. 255-332.
Peña, J. A., Morales, B., y Calderón, R., 1997, New method for comparing levels of microdifferentiation: Application to migration matrices of two populations from the Basque Country (Spain). Human Biology, 69, 329-344.
Roberts, D. F., 1982, Population structure of farming communities of Northern England. En Current developments in Anthropological Genetics. Volume 2: Ecology and population structure, editado por M. Crawford y J. Mielke (New York: Plenum Press) pp. 367-384.
Workman, P., Harpending, H., Lalouel, J., Lynch, C., Niswander, J., y Singleton, R., 1973, Population studies on Southwestern Indian tribes. VI. Papago population structure: a comparison of genetic and migration analyses. En Genetic Structure of Populations, editado por N. Morton (Honolulu: University of Hawaii Press) pp: 166-194.